

IFITM3 requires an amphipathic helix for antiviral activity
- PMID: 28835547
- PMCID: PMC5623871
- DOI: 10.15252/embr.201744100
IFITM3 requires an amphipathic helix for antiviral activity
Abstract
Interferon-induced transmembrane protein 3 (IFITM3) is a cellular factor that blocks virus fusion with cell membranes. IFITM3 has been suggested to alter membrane curvature and fluidity, though its exact mechanism of action is unclear. Using a bioinformatic approach, we predict IFITM3 secondary structures and identify a highly conserved, short amphipathic helix within a hydrophobic region of IFITM3 previously thought to be a transmembrane domain. Consistent with the known ability of amphipathic helices to alter membrane properties, we show that this helix and its amphipathicity are required for the IFITM3-dependent inhibition of influenza virus, Zika virus, vesicular stomatitis virus, Ebola virus, and human immunodeficiency virus infections. The homologous amphipathic helix within IFITM1 is also required for the inhibition of infection, indicating that IFITM proteins possess a conserved mechanism of antiviral action. We further demonstrate that the amphipathic helix of IFITM3 is required to block influenza virus hemagglutinin-mediated membrane fusion. Overall, our results provide evidence that IFITM proteins utilize an amphipathic helix for inhibiting virus fusion.
Keywords: IFITM; amphipathic helix; fusion; restriction factor; virus.
© 2017 The Authors.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures

- A
Results of secondary structure prediction for the human IFITM3 amino acid sequence by the Iterative Threading Assembly Refinement (I‐TASSER) method. Predicted helical regions are shaded in different colors. The amphipathic helix of interest in this study is shaded orange.
- B, C
Different views of the helix predicted for amino acids 59–68 (VWSLFTNLFM) produced using the PEP‐FOLD prediction program displaying both a hydrophobic (B) and primarily hydrophilic (C) face.
- D
Circular dichroism spectra collected for a synthetic peptide corresponding to IFITM3 amino acids 59–68.
- E
Proposed membrane topology model for IFITM3. Colored regions correspond to predicted helices from (A). S‐Palmitoylation of cysteines 71 and 72 is represented by red lines.
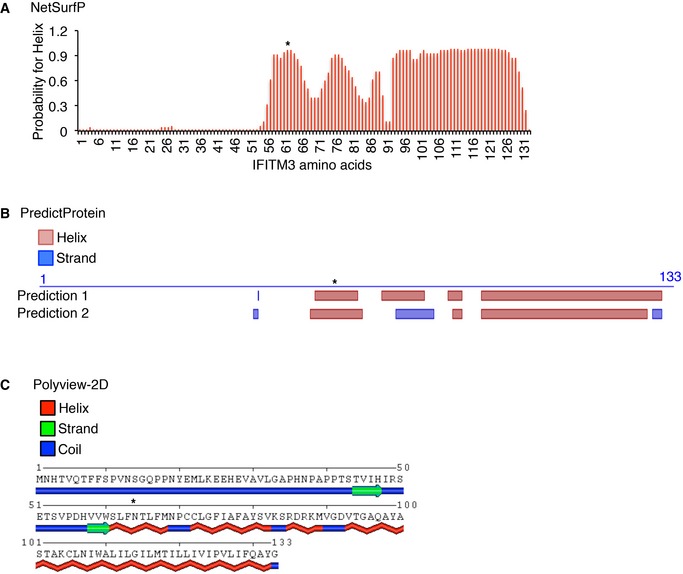
- A–C
Secondary structure predictions for the human IFITM3 amino acid sequence using the (A) NetSurfP, (B) PredictProtein, and (C) POLYVIEW‐2D online programs. *denotes the predicted amphipathic helix region of interest examined in this study.

- A, B
HEK293T cells were transfected overnight with empty vector, full‐length human IFITM3 (WT), IFITM3 lacking the helix at residues 59–68 (Δ59‐68), or palmitoylation‐deficient IFITM3 with cysteines 71, 72, and 105 mutated to alanine (ΔPalm). For (A), cells were infected with H1N1 influenza A virus strain PR8 at an MOI of 2.5 for 6 h and then were fixed, permeabilized, and stained with anti‐IFITM3 to identify transfected cells and with anti‐influenza NP to quantify the percentage of infected cells by flow cytometry. Results shown are an average of n = 3 experiments each performed in triplicate. Error bars represent SD. *P < 0.0001 calculated by Student's t‐test comparing the conditions indicated by brackets. For (B), cell lysates were subjected to Western blotting to confirm comparable expression of IFITM3 (anti‐myc). Anti‐GAPDH served as a loading control.

- A
Visualization of IFITM3 residues 59–68 (VWSLFTNLFM) on a helical wheel projection plot created using HELIQUEST software. The series of mutants generated to test the amphipathicity of this helix is also shown. Hydrophobic residues are displayed as gray or yellow, while hydrophilic residues are displayed as pink or purple. Arrows represent the magnitude and orientation of the mean hydrophobic moment value calculated by HELIQUEST software. Numerical values of the mean hydrophobic moments are shown in Table EV1.
- B–D
HEK293T cells were transfected overnight with empty vector or indicated myc‐IFITM3 mutants. For (B), cells were infected with IAV PR8 at an MOI of 2.5 for 6 h, and then fixed, permeabilized, and stained with anti‐IFITM3 to identify transfected cells and with anti‐influenza NP to quantify the percentage of infected cells by flow cytometry. Averaged results from n = 3 or more experiments, each performed in triplicate, are shown. Error bars represent SEM. **P < 0.0001 and *P < 0.01 in comparison with WT IFITM3 calculated by Student's t‐test. For (C), cell lysates were subjected to Western blotting to determine expression of IFITM3 (anti‐myc) with anti‐GAPDH serving as a control showing comparable loading. For (D), cells were infected with influenza B virus strain B/Texas/06/2011 (IBV) at an MOI of 5 for 24 h, influenza A virus strain X‐31 at an MOI of 5 for 6 h, vesicular stomatitis virus expressing GFP (VSV) at an MOI of 1.0 for 8 h, or Ebola virus Zaire strain glycoprotein‐pseudotyped lentivirus expressing GFP (EBOV) for 48 h. Cells were then fixed, permeabilized, and stained with anti‐myc to identify transfected cells. For IBV and H3N2 infections, cells were also stained with specific anti‐influenza virus nucleoprotein antibodies to quantify the percent infections by flow cytometry. Alternatively, GFP‐positive cells were quantified by flow cytometry for VSV and EBOV infections. Infection percentages were normalized relative to a value of 100% set for vector control infections. Results shown are averages of n = 3 experiments and error bars represent SD.
- E
Flow cytometry histograms of anti‐IFITM3 staining for stable HEK293T and CHME “4 × 4” cell lines transduced with empty lentivirus (vector control), or lentivirus expressing WT IFITM3, IFITM3‐Δ59‐68, or IFITM3‐S61A,N64A,T65A.
- F
HEK293T cell lines as in (E) were infected with H1N1 influenza A virus strain PR8 (H1N1) at an MOI of 0.2 for 18 h or with Zika virus strain HD78 (ZIKV) at an MOI of 1.0 for 72 h. CHME “4 × 4” cell lines as in (E) were infected with 10 ng p24 equivalents of HIV‐1 prepared from molecular clone NL4.3 for 48 h. Infected cells were then stained, respectively, with anti‐influenza virus nucleoprotein, anti‐Zika virus envelope protein, or anti‐HIV‐1 p24 antibodies for quantification of percent infection by flow cytometry. Infection percentages were normalized relative to a value of 100% for vector control infections. Results shown are averages from n = 4 experiments, each performed in triplicate, for each virus, and error bars represent SEM.

Visualization of IFITM3 mutant versions of the IFITM3 residues 59–68 (VWSLFTNLFM) on helical wheel projections created using HELIQUEST software. Hydrophobic residues are displayed as gray or yellow, while hydrophilic residues are displayed as pink or purple. Arrows represent the magnitude and orientation of the mean hydrophobic moment value calculated by HELIQUEST software. Numerical values of the mean hydrophobic moments are shown in Table EV1.
Flow cytometry histograms of anti‐IFITM3 staining for HEK293T cells transfected overnight with the indicated myc‐IFITM3 constructs or vector control showing that each of the mutants was expressed at levels comparable to WT IFITM3.
Cells as in (B) were infected with IAV PR8 at an MOI of 2.5 for 6 h, and then fixed, permeabilized, and stained with anti‐IFITM3 to identify transfected cells, and with anti‐influenza NP to quantify the percentage of infected cells by flow cytometry. Averaged results from n = 3 experiments, each performed in triplicate, are shown. Error bars represent standard deviation. *P < 0.0001 compared to vector control, and **P = 0.009 compared to vector control and **P < 0.0001 compared to WT IFITM3 calculated by Student's t‐test.

- A–C
HeLa cells were transfected for 24 h with the indicated myc‐tagged IFITM3 constructs and (A, B) were fixed, permeabilized, and stained with anti‐myc and anti‐CD63, or (C) were co‐transfected with LAMP1‐GFP and were fixed, permeabilized, and stained with anti‐myc alone. Example fluorescent confocal microscopy images used for colocalization analysis are shown in (A). Manders’ Overlap Coefficient for IFITM3 overlap with CD63 (B) or with LAMP1‐GFP (C) was calculated using the JACoP plugin for ImageJ. For each bar in (B and C), overlap from at least 10 random cell fields from n = 2 experiments, each showing at least 10 cells were quantified and averaged. Error bars represent SEM. No statistical differences were observed between samples as calculated by Student's t‐test.


- A, B
HEK293T cells were transfected overnight with empty vector or the indicated HA‐tagged IFITM1 constructs. For (A), cells were infected with H1N1 influenza A virus strain PR8 at an MOI of 2.5 for 6 h, and then fixed, permeabilized, and stained with anti‐HA to identify transfected cells, and with anti‐influenza NP to quantify the percentage of infected cells by flow cytometry. Results shown are the average of n = 3 experiments, each performed in triplicate. Error bars represent SD. *P < 0.0001 calculated by Student's t‐test comparing the conditions indicated by brackets. For (B), cell lysates were subjected to Western blotting to determine expression of IFITM1 (anti‐HA) with anti‐GAPDH serving as a control showing comparable loading.

HEK293T cells were synchronously infected with H1N1 influenza A virus strain PR8 at an MOI of 40 at 4°C for 1 h to allow virus to bind to cells. Unbound virus was washed away with PBS and cells were collected and fixed (0 h), or the washed cells were incubated in fresh media at 37°C for 1 h followed fixation. Cells collected at 0 and 1 h were stained with the anti‐HA antibody Y8‐10C2 for analysis by flow cytometry. Histograms show representative staining of non‐infected control samples (gray shading) overlayed with infected samples (blue). Results are representative of n = 4 similar experiments.
HEK29T cells stably transduced with IFITM3 or vector control were synchronously infected with H1N1 influenza A virus strain PR8 at an MOI of 40 for 1 h at 4°C. Unbound virus was washed away with PBS, and infection was allowed to proceed for 1 h at 37°C. Cells were trypsinized and washed with PBS to remove extracellular virus. Cells were then fixed, permeabilized, and stained with the indicated anti‐HA antibody clones for analysis by flow cytometry. Descriptions in parentheses indicate the binding specificity of the individual HA antibody clones. Histograms show representative staining of non‐infected control samples (gray shading) overlayed with infected (colored) samples. Bar graphs depict the average mean fluorescence intensity of staining for triplicate infected samples from an experiment representative of n = 3 similar experiments. Error bars represent SD. No statistical differences were observed between samples as calculated by Student's t‐test.

- A
Cell–cell fusion assay schematic.
- B, C
HEK293T cells were transfected with pFR‐Luc plus plasmid expressing (B) IAV HA or (C) VSV G. A second set of cells were transfected with pBD‐NFκB plus plasmid expressing myc‐IFITM3, the indicated myc‐IFITM3 mutants, or vector control. Mixed cell populations were treated with a short, dilute trypsin wash to proteolytically activate HA at the cell surface, immediately followed by a 2‐min incubation with media at pH 5.0 to trigger pH‐dependent cell–cell fusion. After 6 h, cell lysates were analyzed for luciferase activity. Bar graphs depict averages from n = 4 experiments. Error bars represent SD. *P‐value < 0.01 calculated by Student's t‐test in comparison with the vector control.
- D
A proposed model for IFITM3 antiviral activity. In cells lacking IFITM3, the stalk‐pore model 73 for virus fusion posits that influenza virus HA inserts its fusion peptide into the host endosome membrane and further undergoes acid‐induced conformational changes (not shown) that bring virus and host membranes in close proximity (membrane pinching). Mixing of the contacting membranes forms a stalk intermediate, which transitions into a hemifusion diaphragm in which the distal leaflets of the virus and host membranes are in contact, followed by formation of a fusion pore. In the presence of IFITM3 (blue region depicts transmembrane domain, and the orange triangle indicates the membrane wedging mechanism of its amphipathic helix), HA begins the process of virus fusion and proceeds to the hemifusion intermediate 21. We propose that the IFITM3 amphipathic helix is excluded from the hemifusion diaphragm by virtue of its transmembrane domain, but may sense and stabilize curvature produced at the vertexes of the diaphragm, thus preventing its expansion and rupture into a fusion pore.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
