

Src kinase inhibition reduces inflammatory and cytoskeletal changes in ΔF508 human cholangiocytes and improves cystic fibrosis transmembrane conductance regulator correctors efficacy
- PMID: 28836688
- PMCID: PMC5783790
- DOI: 10.1002/hep.29400
Src kinase inhibition reduces inflammatory and cytoskeletal changes in ΔF508 human cholangiocytes and improves cystic fibrosis transmembrane conductance regulator correctors efficacy
Abstract
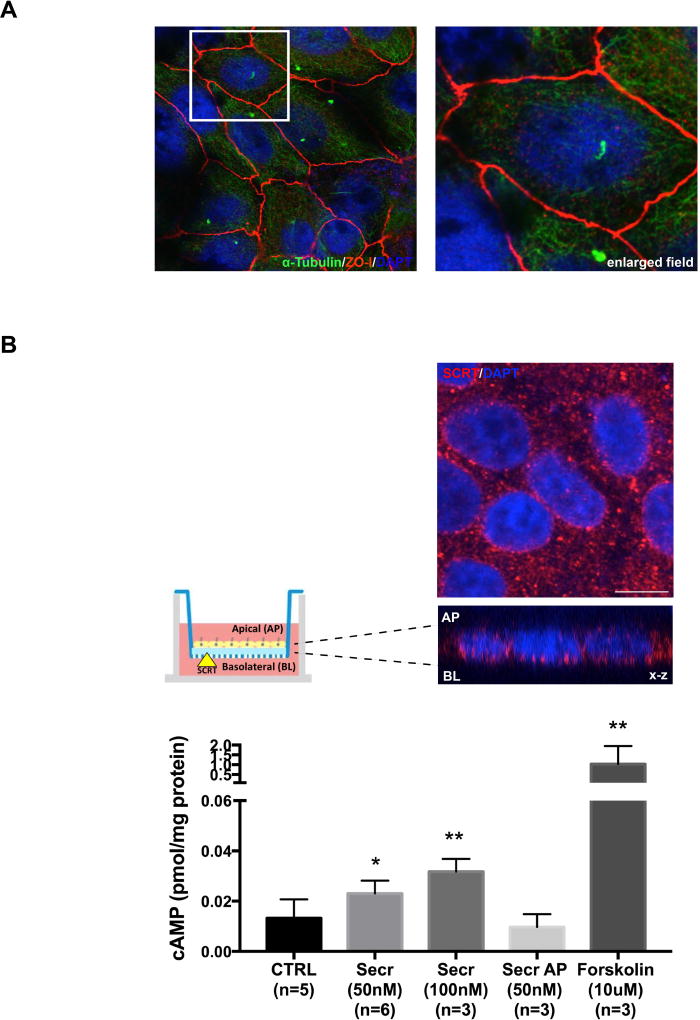
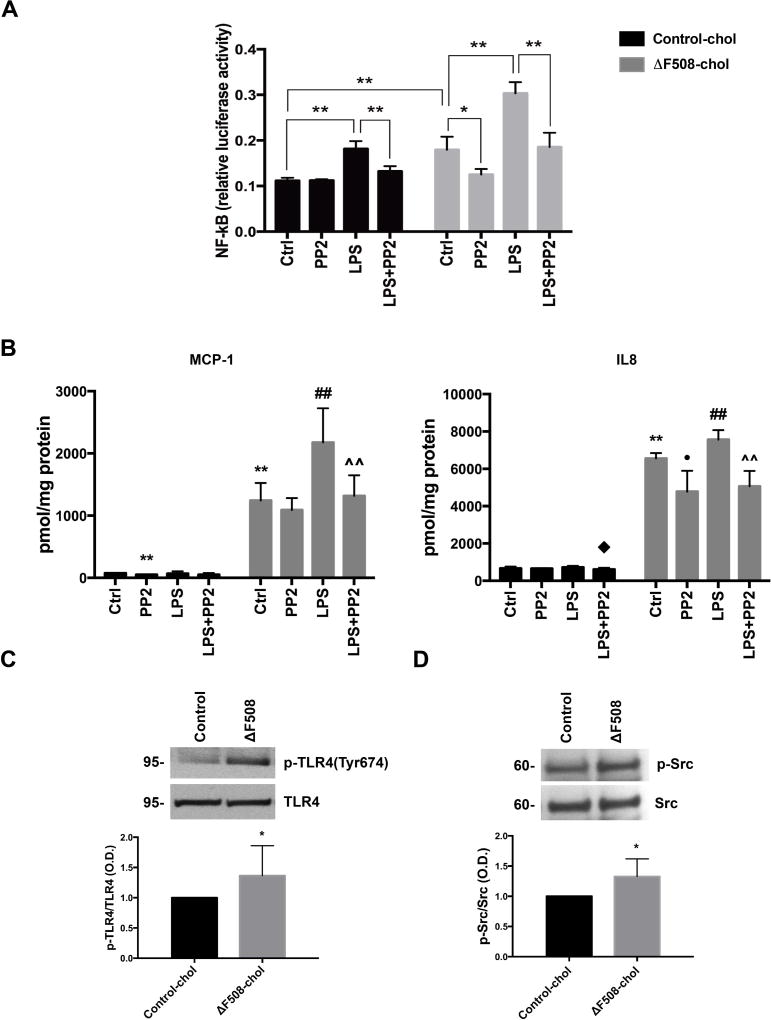
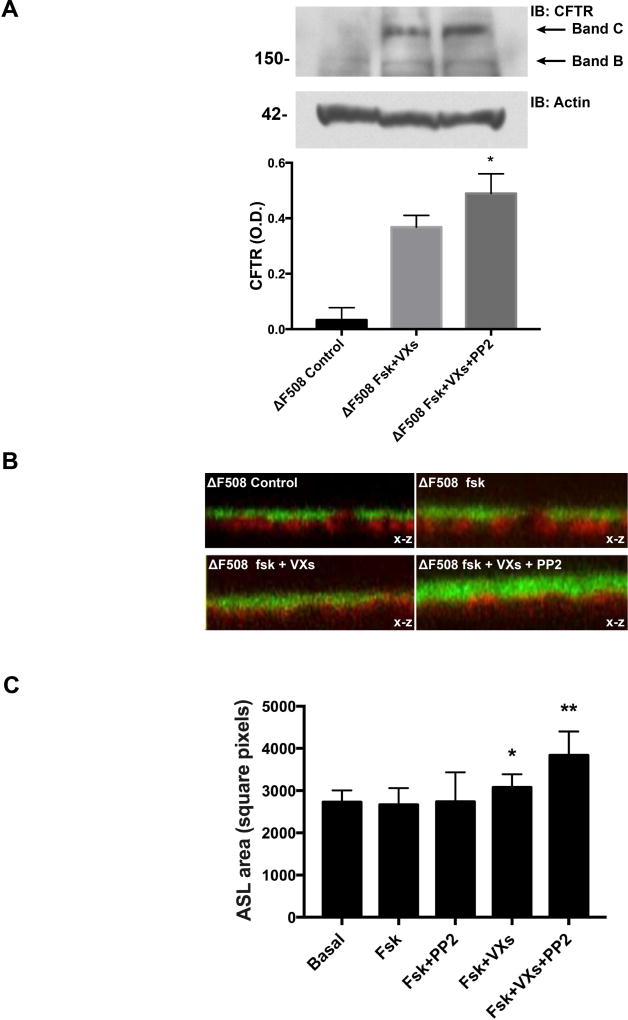
Cystic fibrosis transmembrane conductance regulator (CFTR), the channel mutated in cystic fibrosis (CF), is expressed by the biliary epithelium (i.e., cholangiocytes) of the liver. Progressive clinical liver disease (CF-associated liver disease; CFLD) occurs in around 10% of CF patients and represents the third leading cause of death. Impaired secretion and inflammation contribute to CFLD; however, the lack of human-derived experimental models has hampered the understanding of CFLD pathophysiology and the search for a cure. We have investigated the cellular mechanisms altered in human CF cholangiocytes using induced pluripotent stem cells (iPSCs) derived from healthy controls and a ΔF508 CFTR patient. We have devised a novel protocol for the differentiation of human iPSC into polarized monolayers of cholangiocytes. Our results show that iPSC-cholangiocytes reproduced the polarity and the secretory function of the biliary epithelium. Protein kinase A/cAMP-mediated fluid secretion was impaired in ΔF508 cholangiocytes and negligibly improved by VX-770 and VX-809, two small molecule drugs used to correct and potentiate ΔF508 CFTR. Moreover, ΔF508 cholangiocytes showed increased phosphorylation of Src kinase and Toll-like receptor 4 and proinflammatory changes, including increased nuclear factor kappa-light-chain-enhancer of activated B cells activation, secretion of proinflammatory chemokines (i.e., monocyte chemotactic protein 1 and interleukin-8), as well as alterations of the F-actin cytoskeleton. Treatment with Src inhibitor (4-amino-5-(4-chlorophenyl)-7-(t-butyl)pyrazolo[3,4-d]pyramidine) decreased the inflammatory changes and improved cytoskeletal defects. Inhibition of Src, along with administration of VX-770 and VX-809, successfully restored fluid secretion to normal levels.
Conclusion: Our findings have strong translational potential and indicate that targeting Src kinase and decreasing inflammation may increase the efficacy of pharmacological therapies aimed at correcting the basic ΔF508 defect in CF liver patients. These studies also demonstrate the promise of applying iPSC technology in modeling human cholangiopathies. (Hepatology 2018;67:972-988).
© 2017 by the American Association for the Study of Liver Diseases.
Conflict of interest statement
Figures




Comment in
-
Cystic fibrosis transmembrane conductance regulator: Actin(g) as a master regulator of cholangiocyte function.Hepatology. 2018 Mar;67(3):833-836. doi: 10.1002/hep.29583. Epub 2018 Jan 26. Hepatology. 2018. PMID: 29023826 No abstract available.
References
-
- Riordan JR, Rommens JM, Kerem B, Alon N, Rozmahel R, Grzelczak Z, et al. Identification of the cystic fibrosis gene: cloning and characterization of complementary DNA. Science. 1989;245:1066–1073. - PubMed
-
- Kinnman N, Lindblad A, Housset C, Buentke E, Scheynius A, Strandvik B, et al. Expression of cystic fibrosis transmembrane conductance regulator in liver tissue from patients with cystic fibrosis. Hepatology. 2000;32:334–340. - PubMed
-
- Rowe SM, Miller S, Sorscher EJ. Cystic fibrosis. N Engl J Med. 2005;352:1992–2001. - PubMed
-
- Colombo C, Battezzati PM, Crosignani A, Morabito A, Costantini D, Padoan R, et al. Liver disease in cystic fibrosis: A prospective study on incidence, risk factors, and outcome. Hepatology. 2002;36:1374–1382. - PubMed
-
- Lindblad A, Glaumann H, Strandvik B. Natural history of liver disease in cystic fibrosis. Hepatology. 1999;30:1151–1158. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
