

Green fluorescent genetically encoded calcium indicator based on calmodulin/M13-peptide from fungi
- PMID: 28837632
- PMCID: PMC5570312
- DOI: 10.1371/journal.pone.0183757
Green fluorescent genetically encoded calcium indicator based on calmodulin/M13-peptide from fungi
Abstract
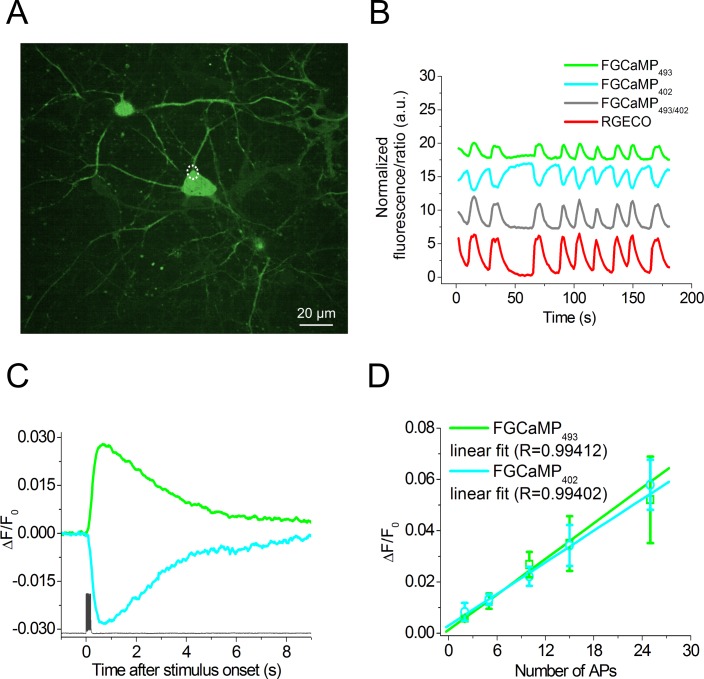
Currently available genetically encoded calcium indicators (GECIs) utilize calmodulins (CaMs) or troponin C from metazoa such as mammals, birds, and teleosts, as calcium-binding domains. The amino acid sequences of the metazoan calcium-binding domains are highly conserved, which may limit the range of the GECI key parameters and cause undesired interactions with the intracellular environment in mammalian cells. Here we have used fungi, evolutionary distinct organisms, to derive CaM and its binding partner domains and design new GECI with improved properties. We applied iterative rounds of molecular evolution to develop FGCaMP, a novel green calcium indicator. It includes the circularly permuted version of the enhanced green fluorescent protein (EGFP) sandwiched between the fungal CaM and a fragment of CaM-dependent kinase. FGCaMP is an excitation-ratiometric indicator that has a positive and an inverted fluorescence response to calcium ions when excited at 488 and 405 nm, respectively. Compared with the GCaMP6s indicator in vitro, FGCaMP has a similar brightness at 488 nm excitation, 7-fold higher brightness at 405 nm excitation, and 1.3-fold faster calcium ion dissociation kinetics. Using site-directed mutagenesis, we generated variants of FGCaMP with improved binding affinity to calcium ions and increased the magnitude of FGCaMP fluorescence response to low calcium ion concentrations. Using FGCaMP, we have successfully visualized calcium transients in cultured mammalian cells. In contrast to the limited mobility of GCaMP6s and G-GECO1.2 indicators, FGCaMP exhibits practically 100% molecular mobility at physiological concentrations of calcium ion in mammalian cells, as determined by photobleaching experiments with fluorescence recovery. We have successfully monitored the calcium dynamics during spontaneous activity of neuronal cultures using FGCaMP and utilized whole-cell patch clamp recordings to further characterize its behavior in neurons. Finally, we used FGCaMP in vivo to perform structural and functional imaging of zebrafish using wide-field, confocal, and light-sheet microscopy.
Conflict of interest statement
Figures
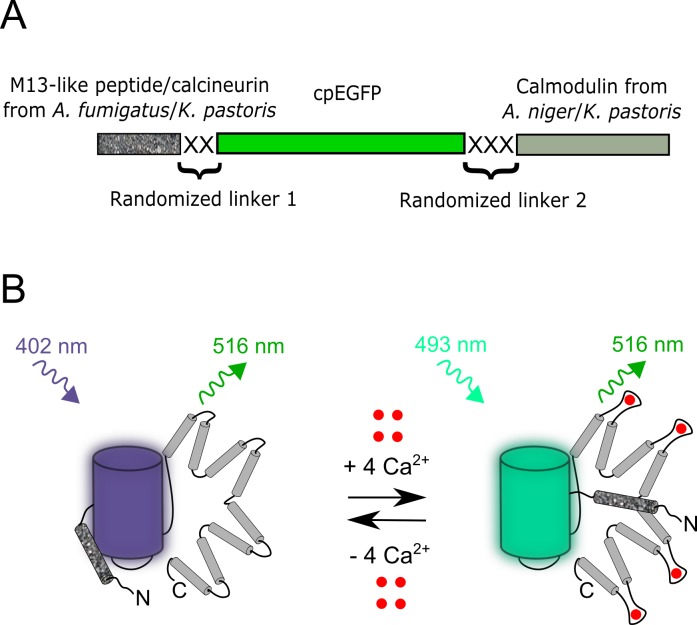
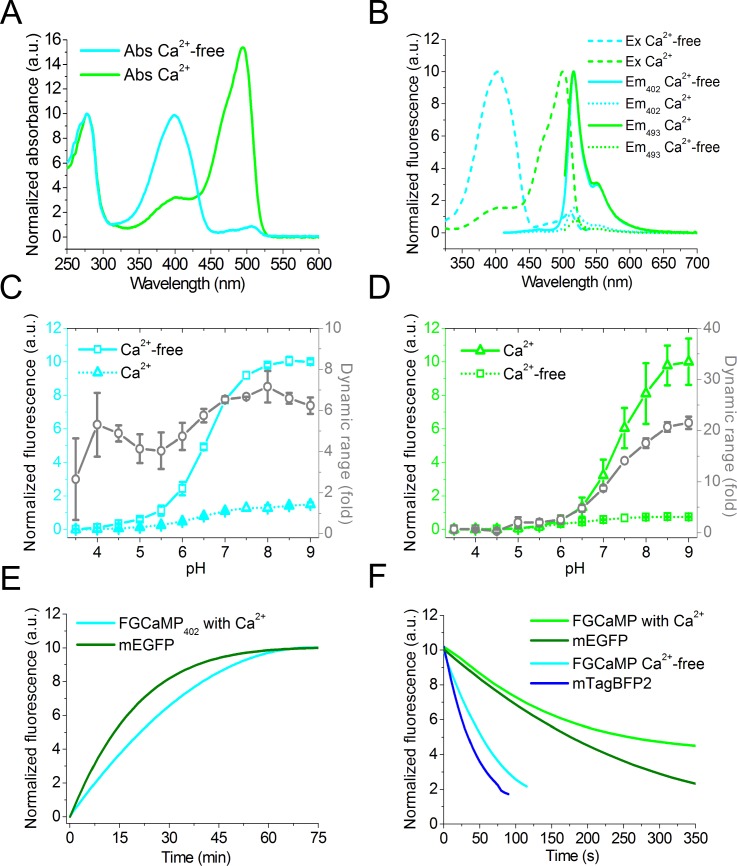
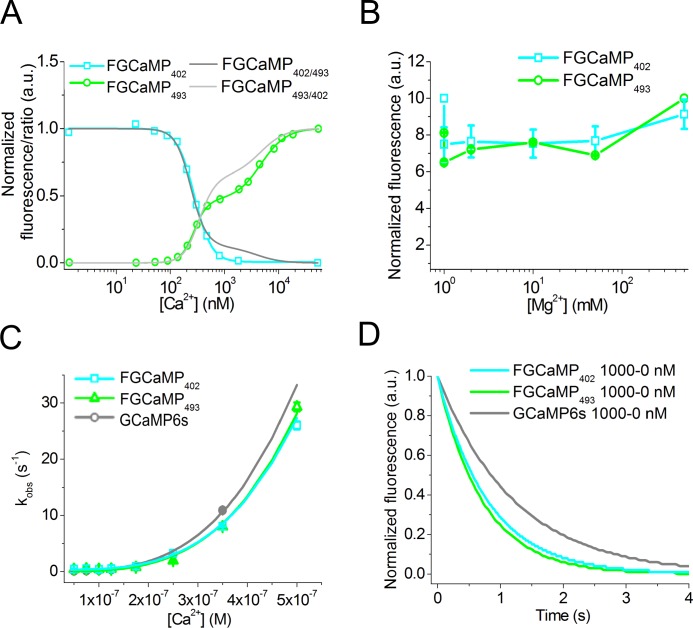
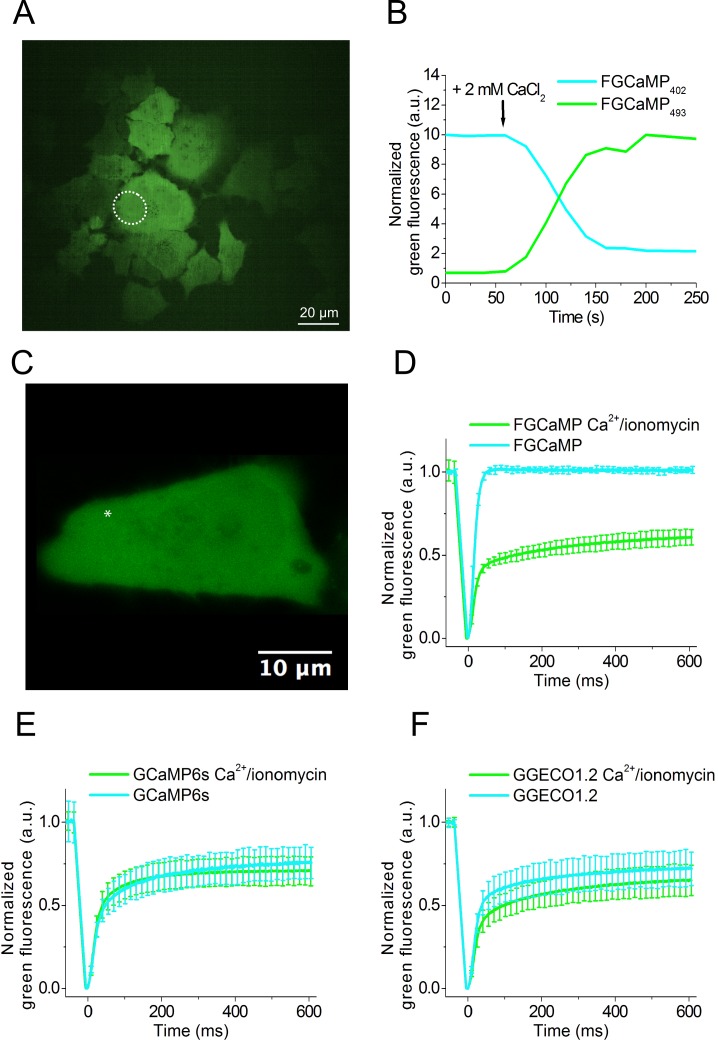

References
-
- Akerboom J, Rivera JD, Guilbe MM, Malave EC, Hernandez HH, Tian L, et al. (2009) Crystal structures of the GCaMP calcium sensor reveal the mechanism of fluorescence signal change and aid rational design. The Journal of biological chemistry 284: 6455–6464. doi: 10.1074/jbc.M807657200 - DOI - PMC - PubMed
-
- Wang Q, Shui B, Kotlikoff MI, Sondermann H (2008) Structural basis for calcium sensing by GCaMP2. Structure 16: 1817–1827. doi: 10.1016/j.str.2008.10.008 - DOI - PMC - PubMed
-
- Griesbeck O, Baird GS, Campbell RE, Zacharias DA, Tsien RY (2001) Reducing the environmental sensitivity of yellow fluorescent protein. Mechanism and applications. The Journal of biological chemistry 276: 29188–29194. doi: 10.1074/jbc.M102815200 - DOI - PubMed
-
- Barykina NV, Subach OM, Doronin DA, Sotskov VP, Roshchina MA, Kunitsyna TA, et al. (2016) A new design for a green calcium indicator with a smaller size and a reduced number of calcium-binding sites. Scientific reports 6: 34447 doi: 10.1038/srep34447 - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
