

Quantitative tests of a reconstitution model for RNA folding thermodynamics and kinetics
- PMID: 28839094
- PMCID: PMC5604005
- DOI: 10.1073/pnas.1703507114
Quantitative tests of a reconstitution model for RNA folding thermodynamics and kinetics
Abstract
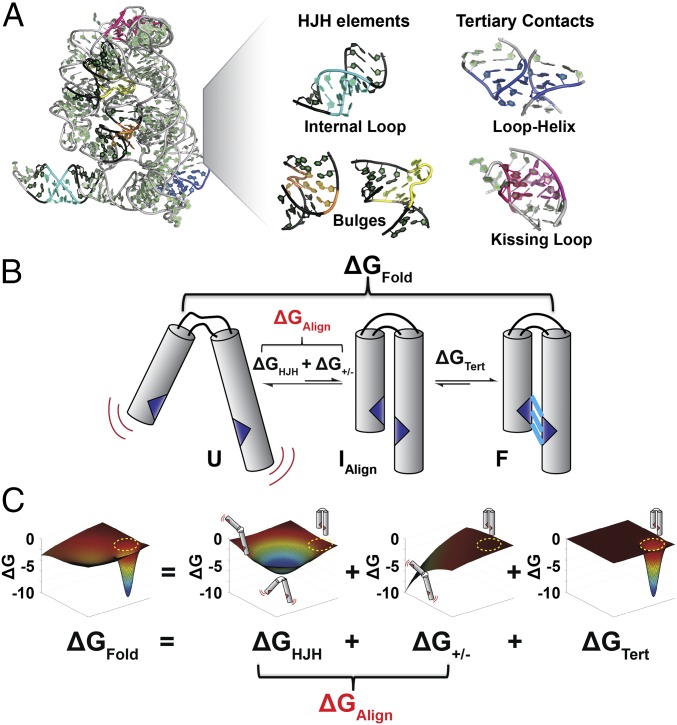
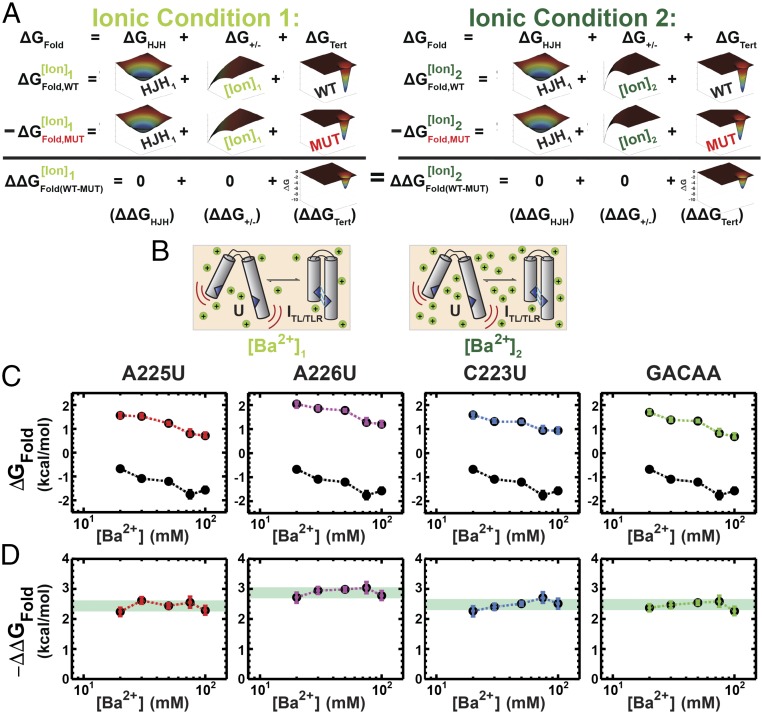
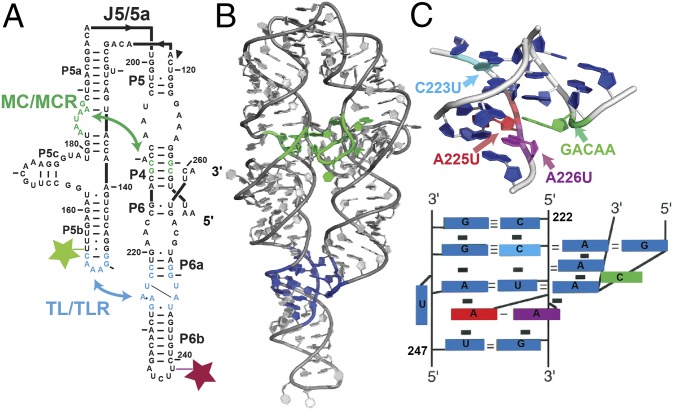
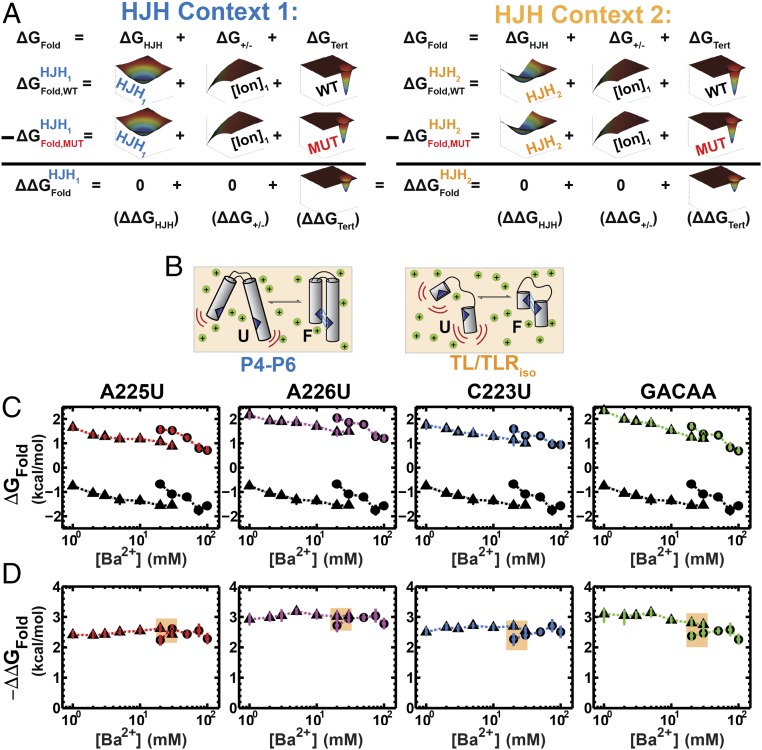
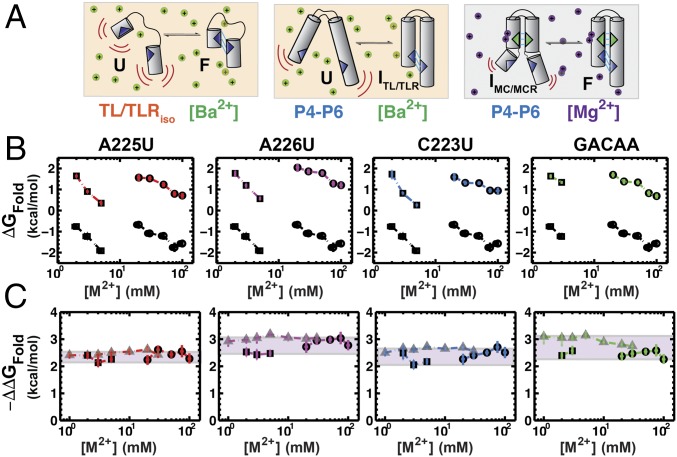
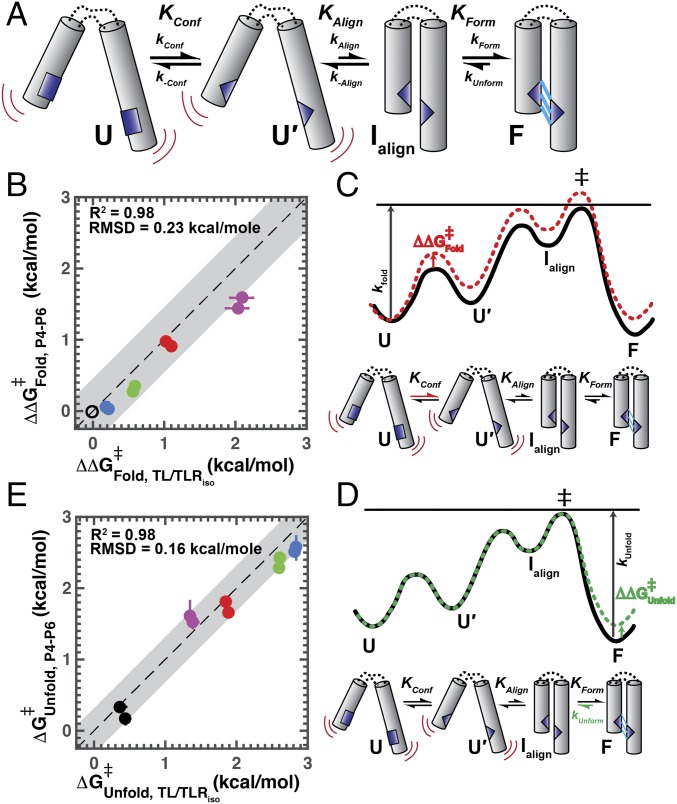
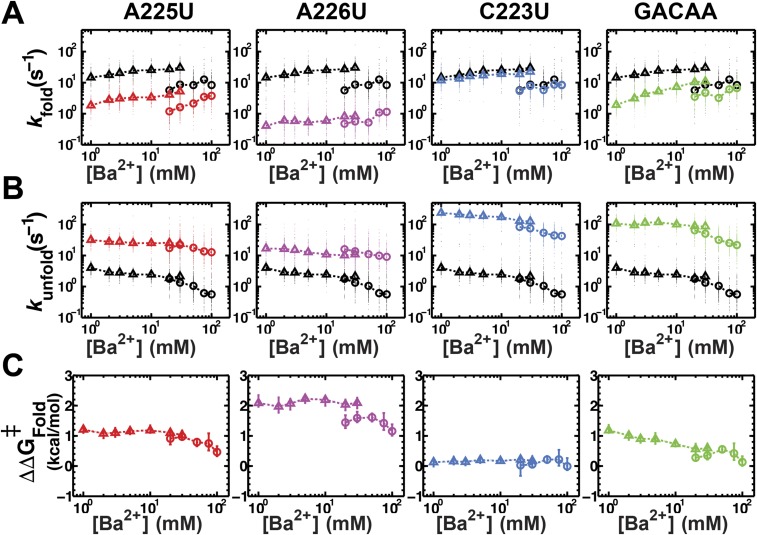
Decades of study of the architecture and function of structured RNAs have led to the perspective that RNA tertiary structure is modular, made of locally stable domains that retain their structure across RNAs. We formalize a hypothesis inspired by this modularity-that RNA folding thermodynamics and kinetics can be quantitatively predicted from separable energetic contributions of the individual components of a complex RNA. This reconstitution hypothesis considers RNA tertiary folding in terms of ΔGalign, the probability of aligning tertiary contact partners, and ΔGtert, the favorable energetic contribution from the formation of tertiary contacts in an aligned state. This hypothesis predicts that changes in the alignment of tertiary contacts from different connecting helices and junctions (ΔGHJH) or from changes in the electrostatic environment (ΔG+/-) will not affect the energetic perturbation from a mutation in a tertiary contact (ΔΔGtert). Consistent with these predictions, single-molecule FRET measurements of folding of model RNAs revealed constant ΔΔGtert values for mutations in a tertiary contact embedded in different structural contexts and under different electrostatic conditions. The kinetic effects of these mutations provide further support for modular behavior of RNA elements and suggest that tertiary mutations may be used to identify rate-limiting steps and dissect folding and assembly pathways for complex RNAs. Overall, our model and results are foundational for a predictive understanding of RNA folding that will allow manipulation of RNA folding thermodynamics and kinetics. Conversely, the approaches herein can identify cases where an independent, additive model cannot be applied and so require additional investigation.
Keywords: RNA folding; RNA tertiary structure; folding kinetics; folding thermodynamics; single-molecule FRET.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Cech TR, Steitz JA. The noncoding RNA revolution-trashing old rules to forge new ones. Cell. 2014;157:77–94. - PubMed
-
- Moore MJ. From birth to death: The complex lives of eukaryotic mRNAs. Science. 2005;309:1514–1518. - PubMed
-
- Staley JP, Guthrie C. Mechanical devices of the spliceosome: Motors, clocks, springs, and things. Cell. 1998;92:315–326. - PubMed
-
- Noller HF. RNA structure: Reading the ribosome. Science. 2005;309:1508–1514. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
