

Combinatorial bioactive botanicals re-sensitize tamoxifen treatment in ER-negative breast cancer via epigenetic reactivation of ERα expression
- PMID: 28839265
- PMCID: PMC5570897
- DOI: 10.1038/s41598-017-09764-3
Combinatorial bioactive botanicals re-sensitize tamoxifen treatment in ER-negative breast cancer via epigenetic reactivation of ERα expression
Abstract
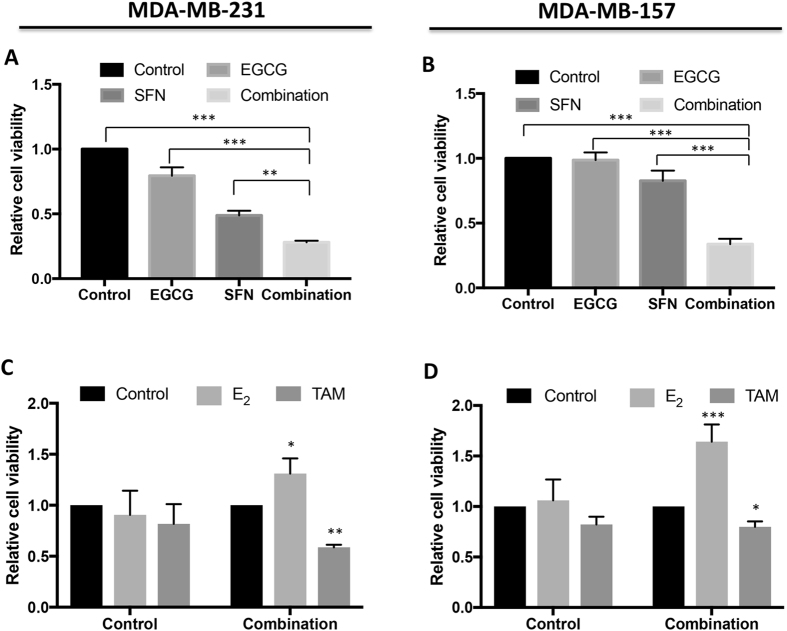
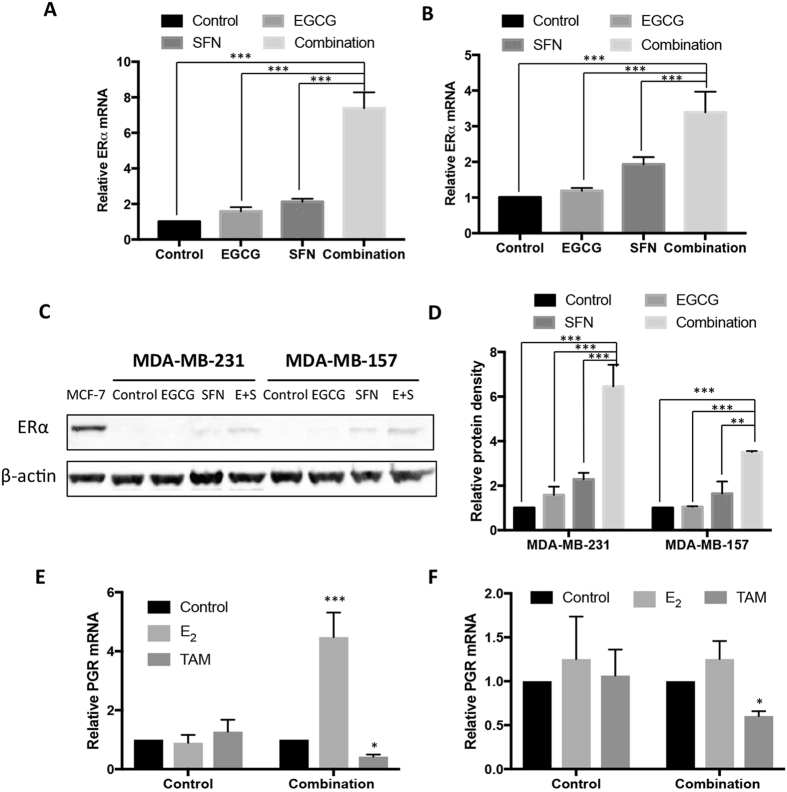
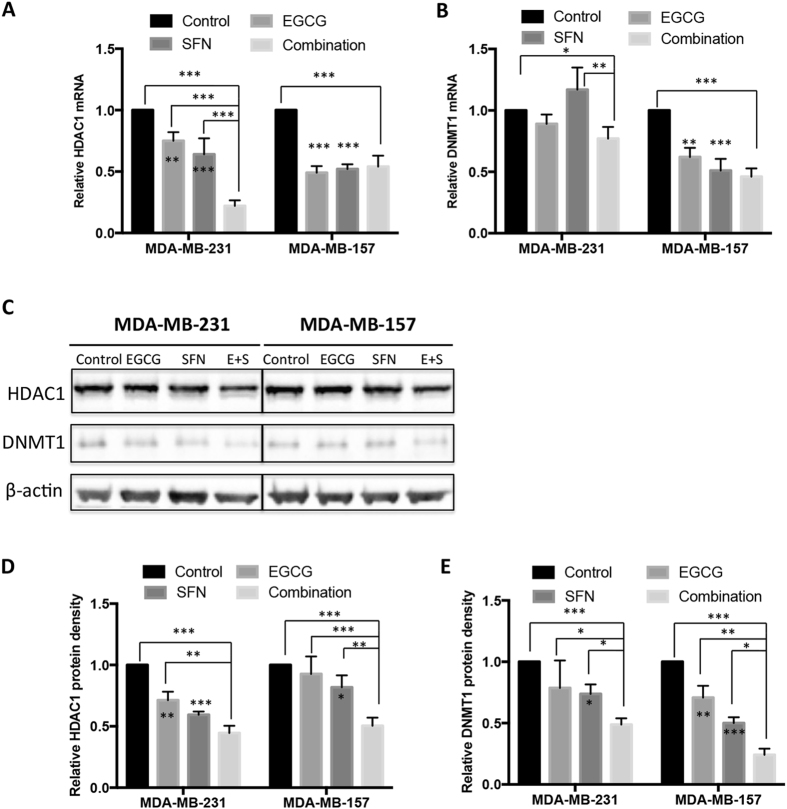
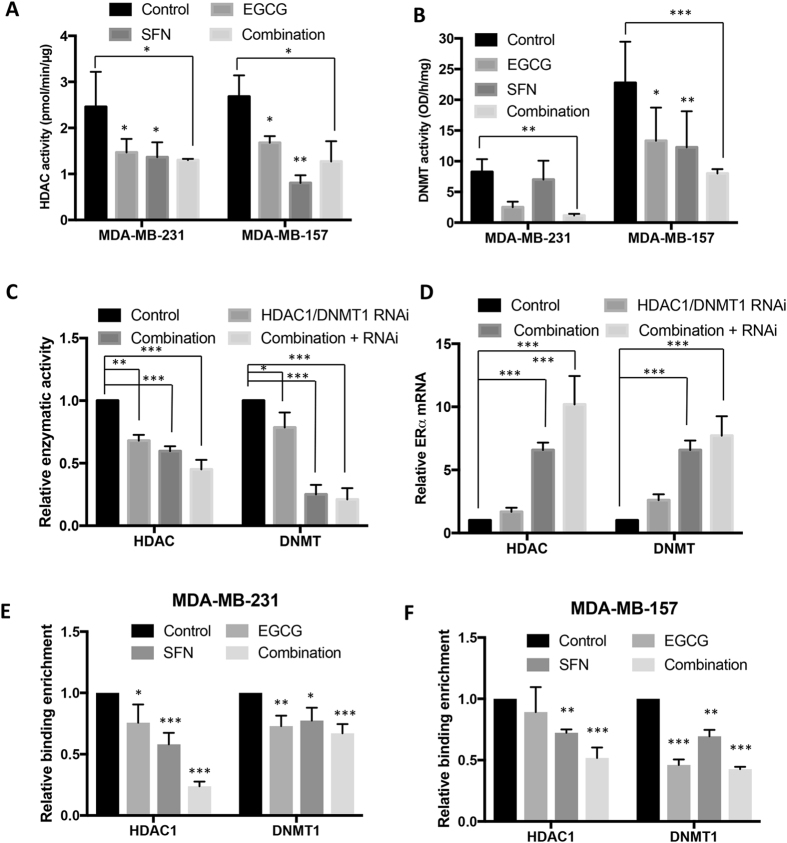
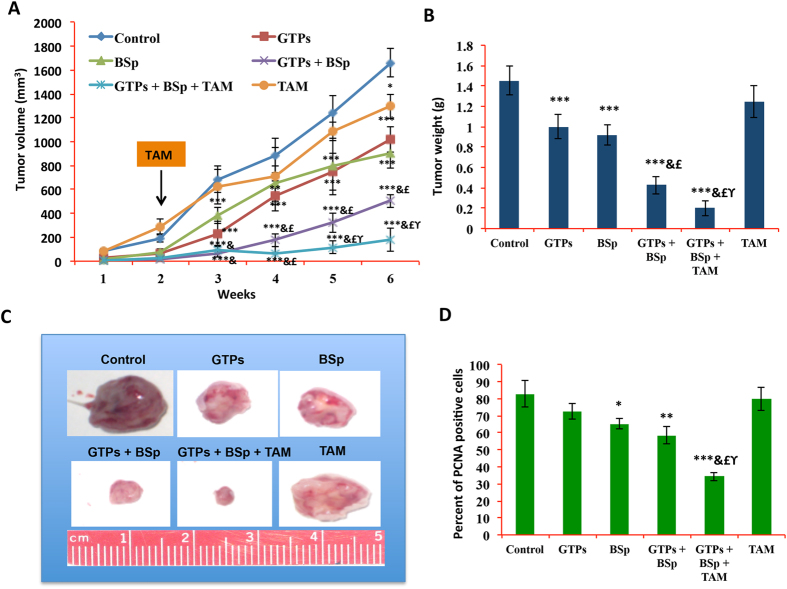
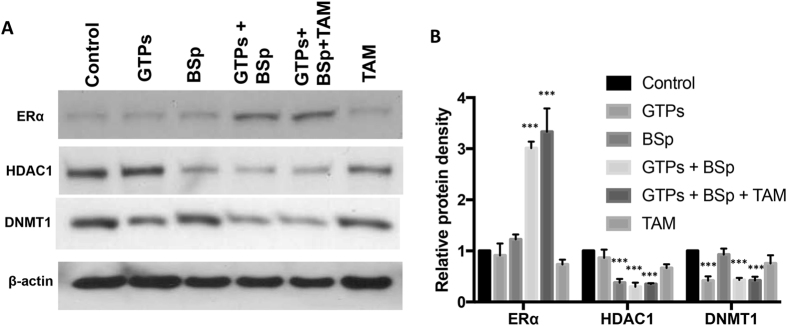
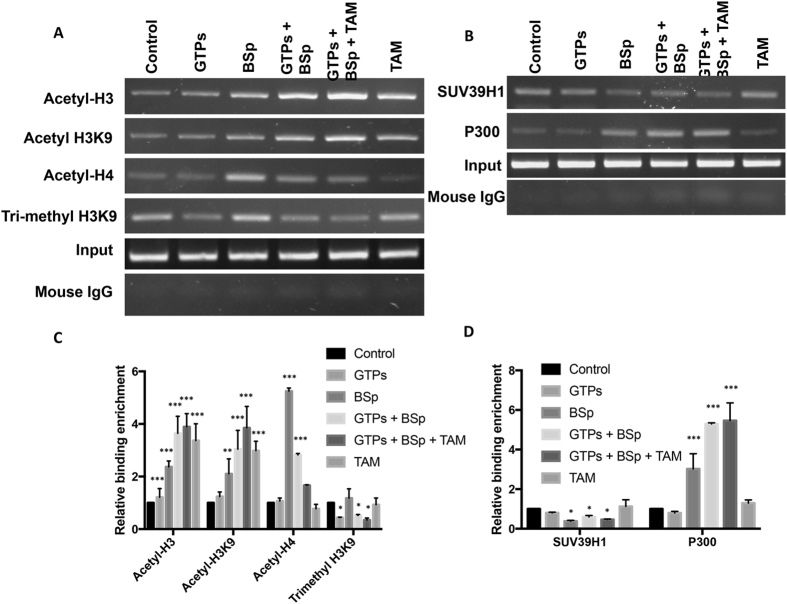
Conventional cancer prevention has primarily focused on single chemopreventive compounds that may not be sufficiently efficacious. We sought to investigate potential combinatorial effects of epigenetic bioactive botanicals including epigallocatechin-3-gallate (EGCG) in green tea polyphenols (GTPs) and sulforaphane (SFN) in broccoli sprouts (BSp) on neutralizing epigenetic aberrations in estrogen receptor-α (ERα) leading to enhanced anti-hormone therapeutic efficacy in ERα-negative breast cancer. Our results showed that this combinatorial treatment re-sensitized ERα-dependent cellular inhibitory responses to an estrogen antagonist, tamoxifen (TAM), via at least in part, epigenetic reactivation of ERα expression in ERα-negative breast cancer cells. Further in vivo studies revealed the combinatorial diets of GTPs and BSp significantly inhibited breast tumor growth in ERα-negative mouse xenografts, especially when combined with TAM treatment. This novel treatment regimen can lead to remodeling of the chromatin structure by histone modifications and recruitment changes of transcriptional factor complex in the ERα promoter thereby contributing to ERα reactivation and re-sensitized chemotherapeutic efficacy of anti-hormone therapy. Our studies indicate that combinatorial bioactive botanicals from GTPs and BSp are highly effective in inhibiting ERα-negative breast cancer due at least in part to epigenetic reactivation of ERα, which in turn increases TAM-dependent anti-estrogen chemosensitivity in vitro and in vivo.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
