

Two CYP82D Enzymes Function as Flavone Hydroxylases in the Biosynthesis of Root-Specific 4'-Deoxyflavones in Scutellaria baicalensis
- PMID: 28842248
- PMCID: PMC5770198
- DOI: 10.1016/j.molp.2017.08.009
Two CYP82D Enzymes Function as Flavone Hydroxylases in the Biosynthesis of Root-Specific 4'-Deoxyflavones in Scutellaria baicalensis
Abstract
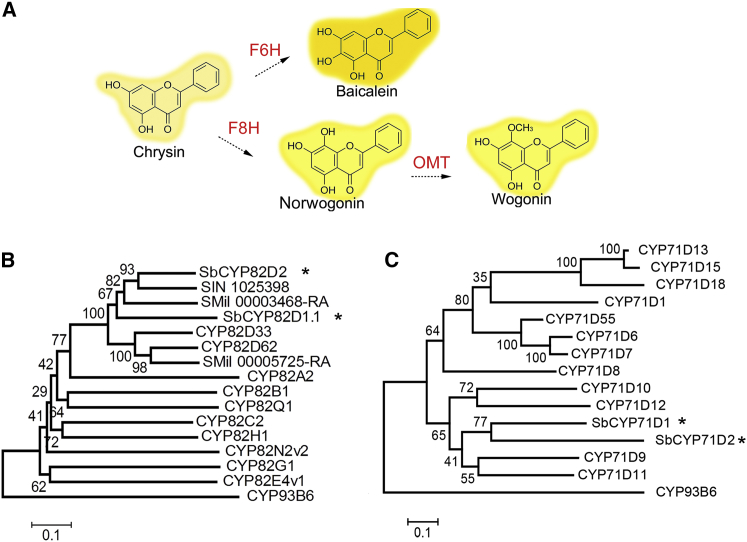
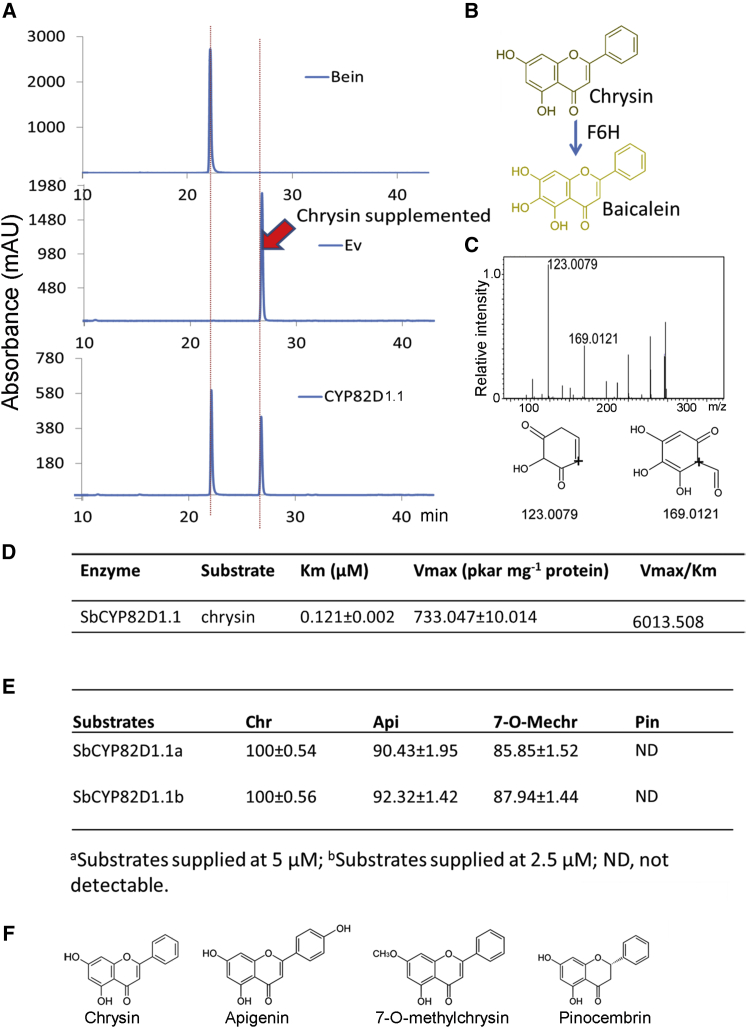
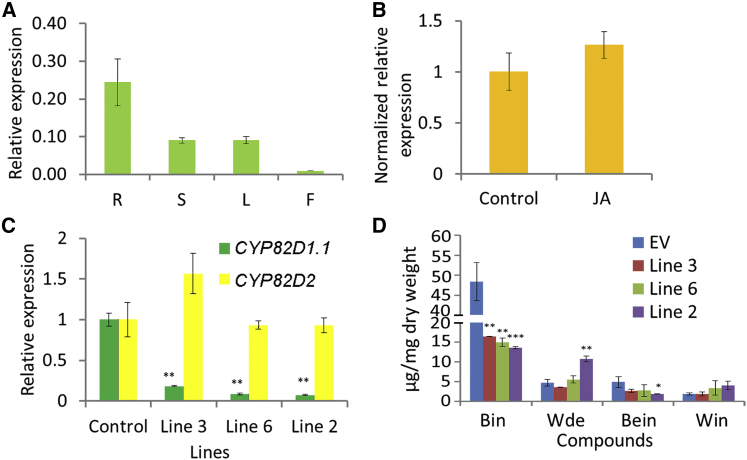
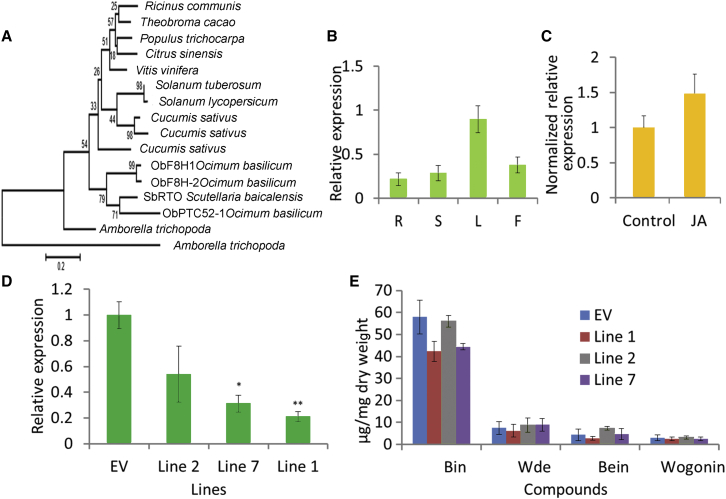
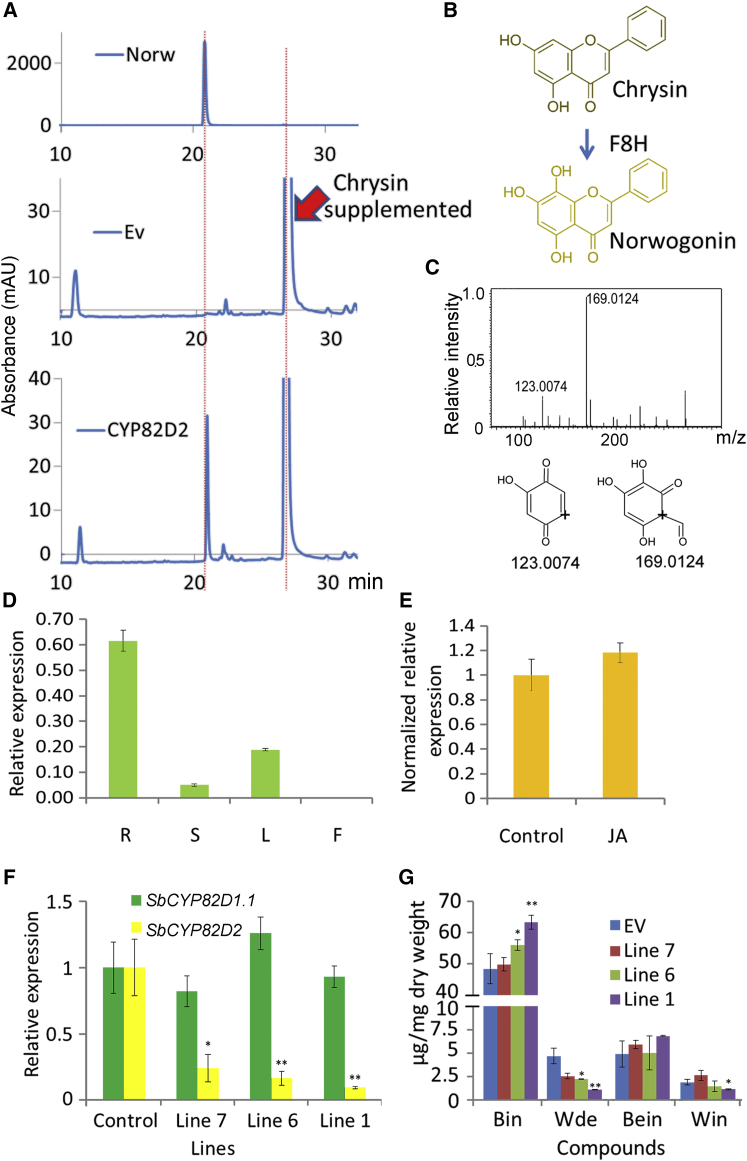
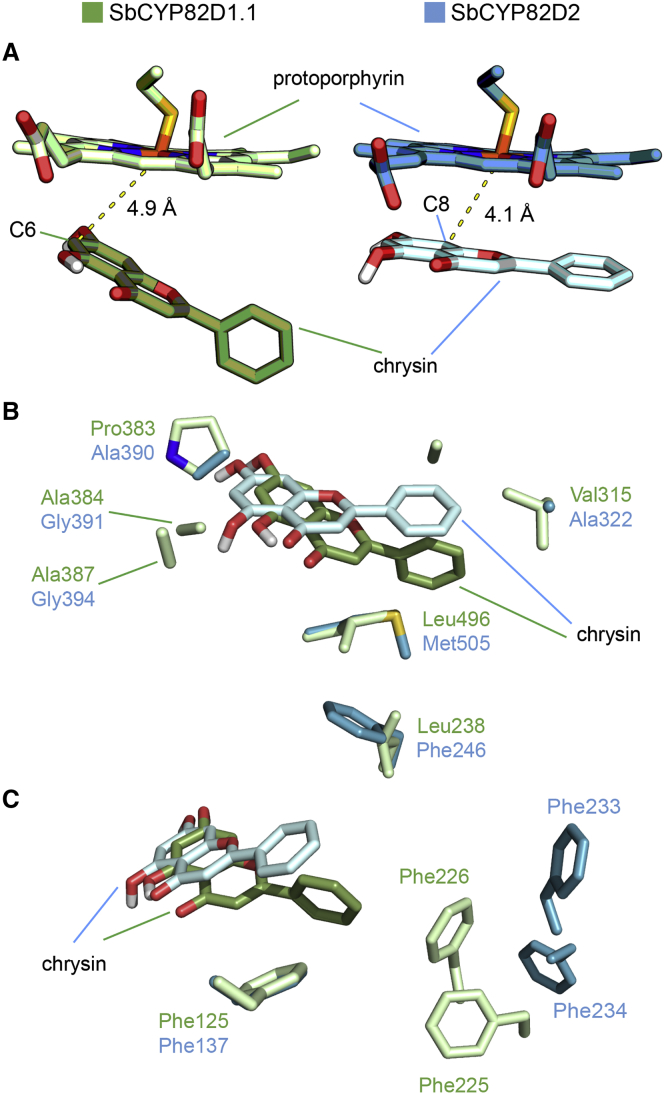
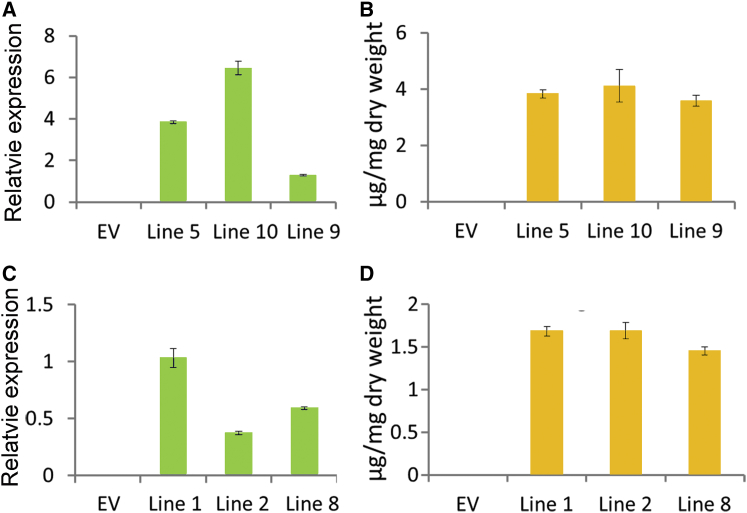
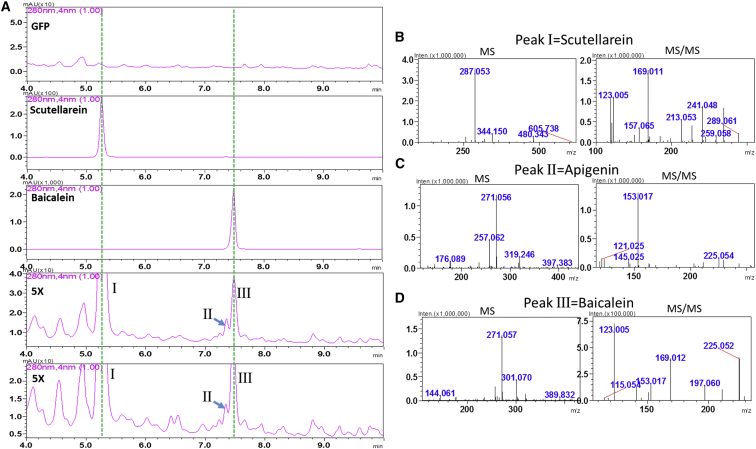
Baicalein, wogonin, and their glycosides are major bioactive compounds found in the medicinal plant Scutellaria baicalensis Georgi. These flavones can induce apoptosis in a variety of cancer cell lines but have no effect on normal cells. Furthermore, they have many additional benefits for human health, such as anti-oxidant, antiviral, and liver-protective properties. Here, we report the isolation and characterization of two CYP450 enzymes, SbCYP82D1.1 and SbCYP82D2, which function as the flavone 6-hydroxylase (F6H) and flavone 8-hydroxylase (F8H), respectively, in S. baicalensis. SbCYP82D1.1 has broad substrate specificity for flavones such as chrysin and apigenin and is responsible for biosynthesis of baicalein and scutellarein in roots and aerial parts of S. baicalensis, respectively. When the expression of SbCYP82D1.1 is knocked down, baicalin and baicalein levels are reduced significantly while chrysin glycosides accumulate in hairy roots. SbCYP82D2 is an F8H with high substrate specificity, accepting only chrysin as its substrate to produce norwogonin, although minor 6-hydroxylation activity can also be detected. Phylogenetic analysis suggested that SbCYP82D2 might have evolved from SbCYP82D1.1 via gene duplication followed by neofunctionalization, whereby the ancestral F6H activity is partially retained in the derived SbCYP82D2.
Keywords: Huangqin; Scutellaria baicalensis; baicalein; flavone 6-hydroxylase; flavone 8-hydroxylase; wogonin.
Copyright © 2017 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Anzellotti D., Ibrahim R.K. Novel flavonol 2-oxoglutarate dependent dioxygenase: affinity purification, characterization, and kinetic properties. Arch. Biochem. Biophys. 2000;382:161–172. - PubMed
-
- Berim A., Park J.J., Gang D.R. Unexpected roles for ancient proteins: flavone 8-hydroxylase in sweet basil trichomes is a Rieske-type, PAO-family oxygenase. Plant J. 2014;80:385–395. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
