

The bacterial condensin MukB compacts DNA by sequestering supercoils and stabilizing topologically isolated loops
- PMID: 28842486
- PMCID: PMC5641887
- DOI: 10.1074/jbc.M117.803312
The bacterial condensin MukB compacts DNA by sequestering supercoils and stabilizing topologically isolated loops
Abstract
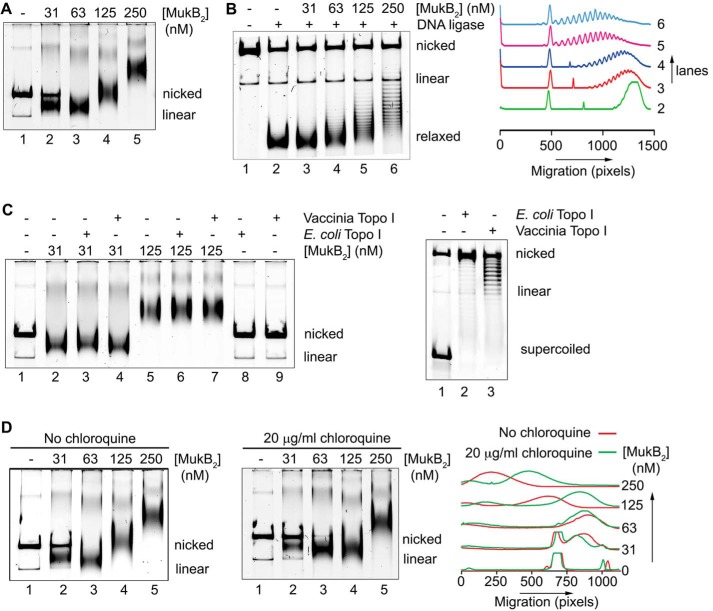
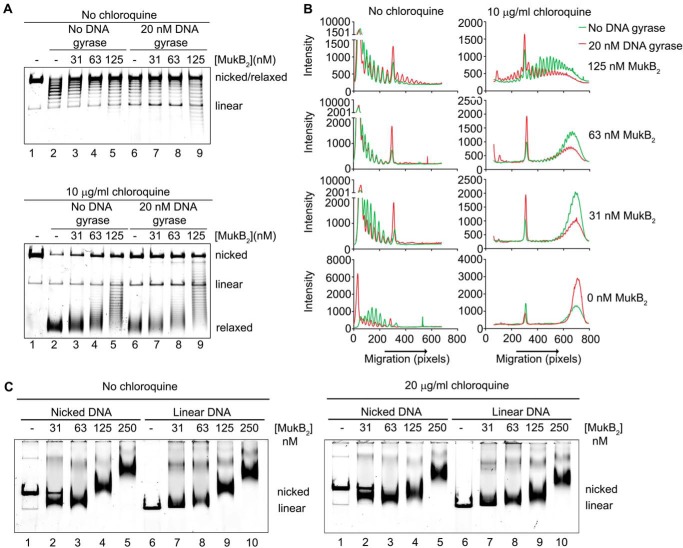
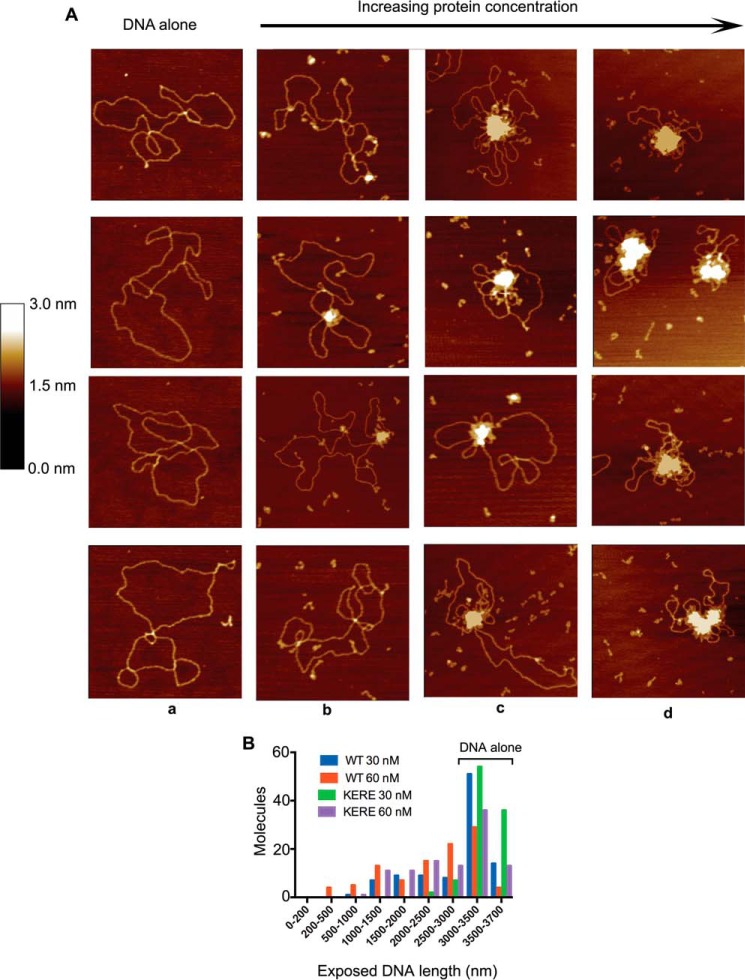
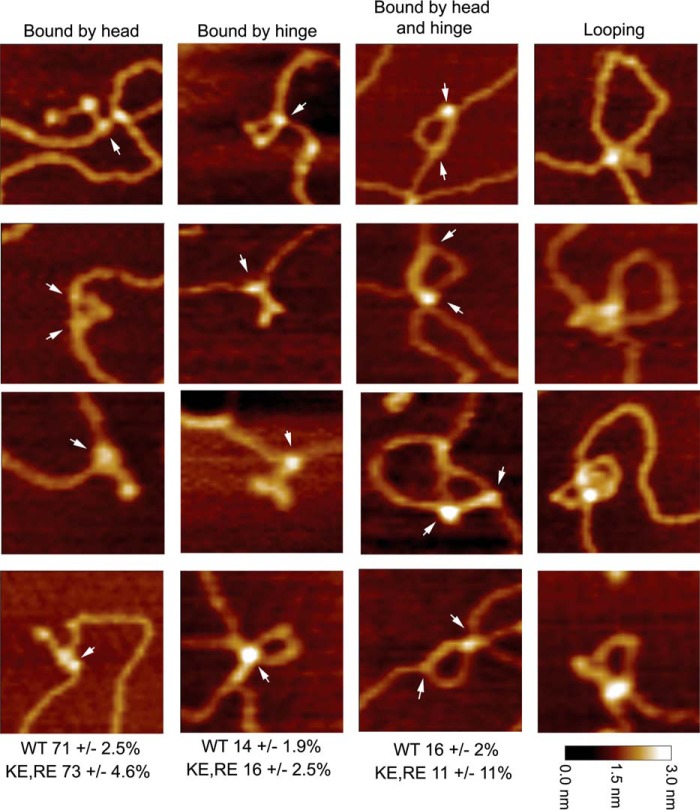
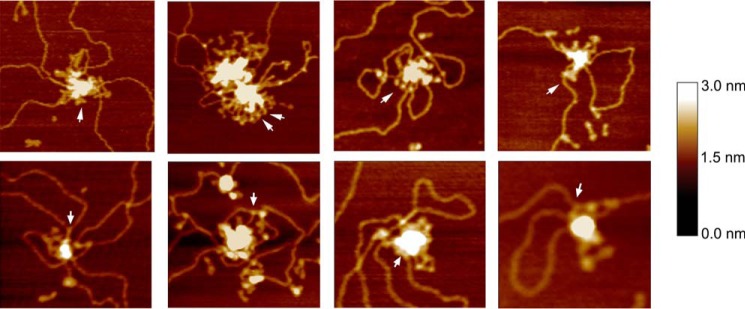
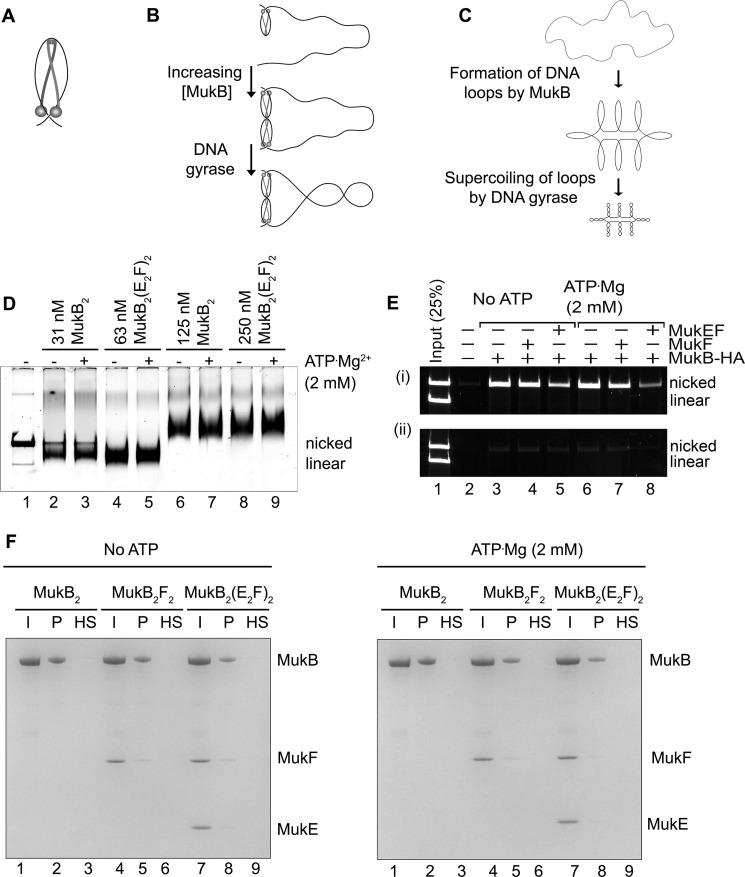
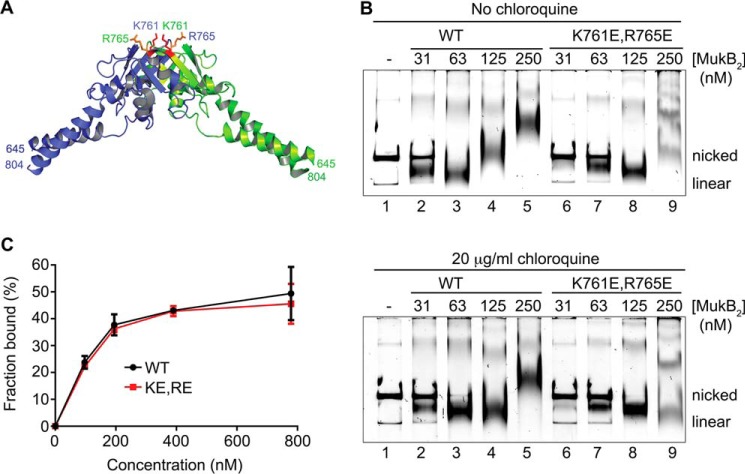
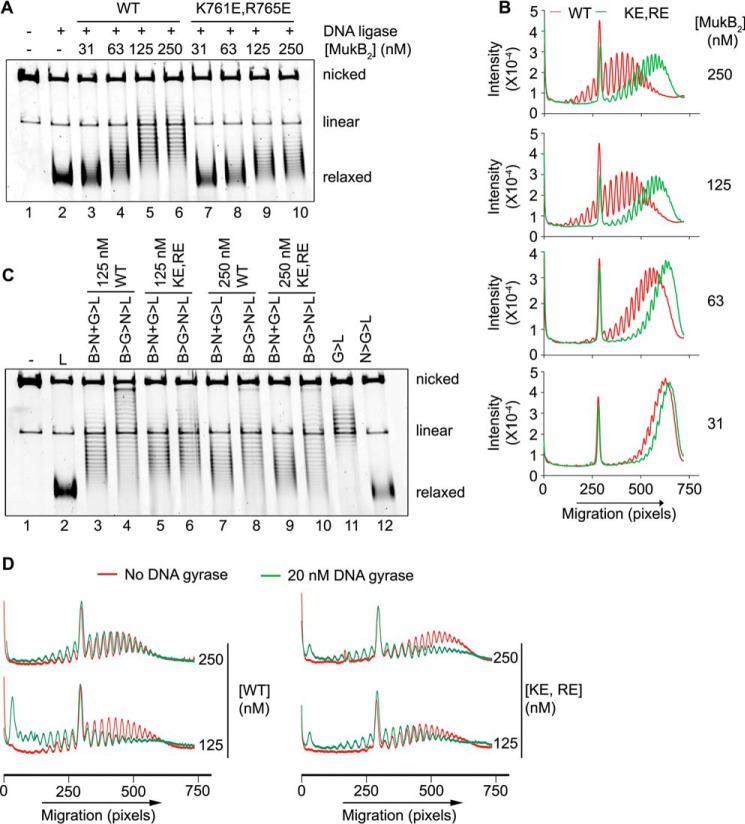
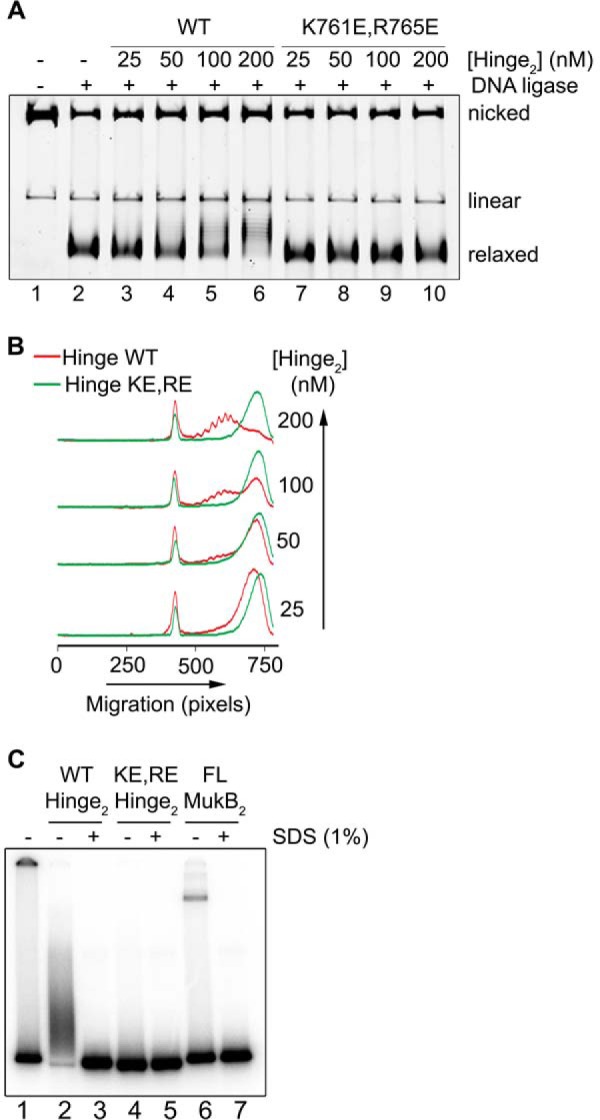
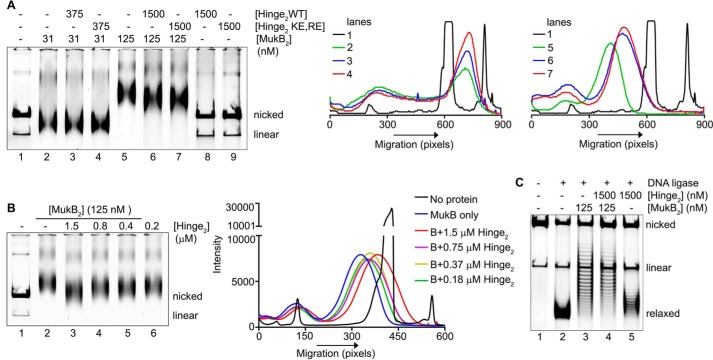
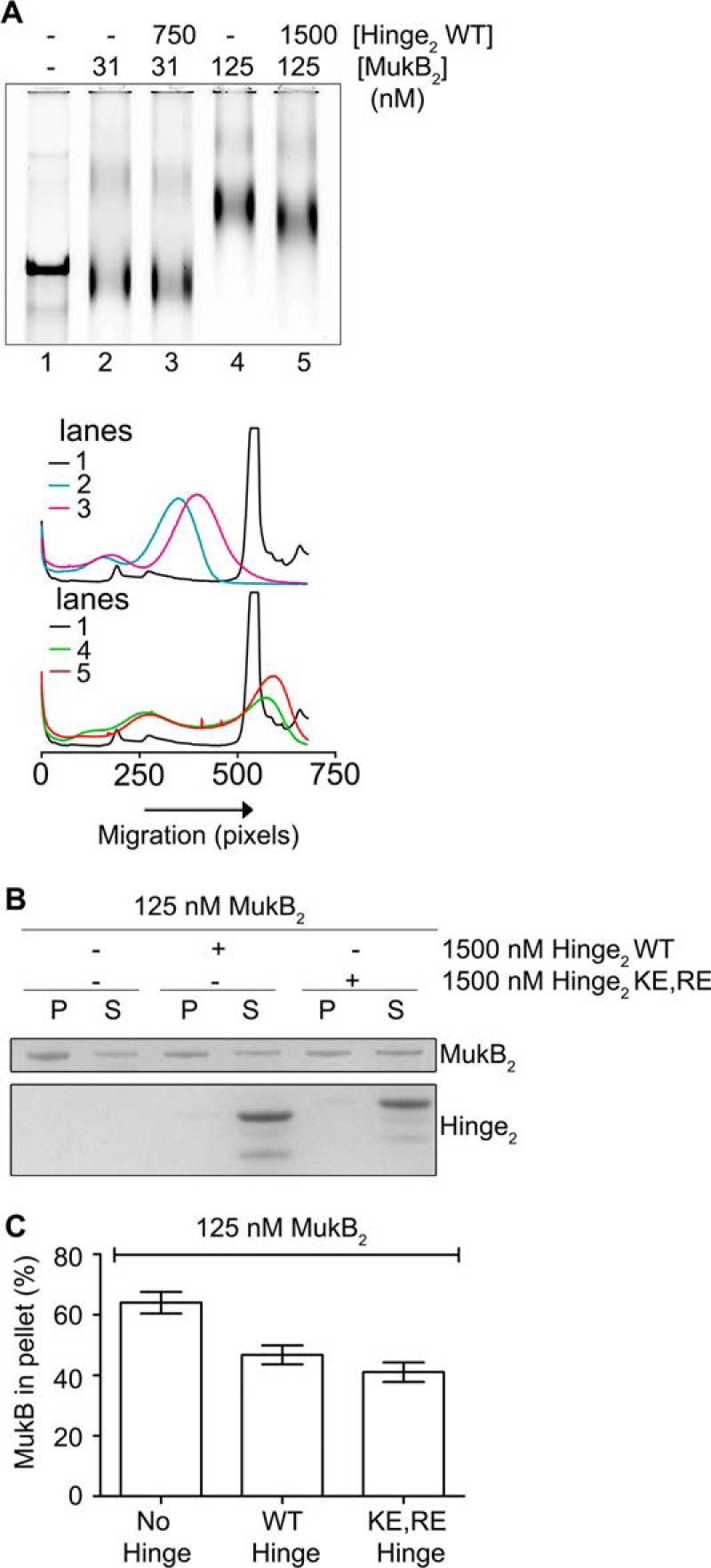
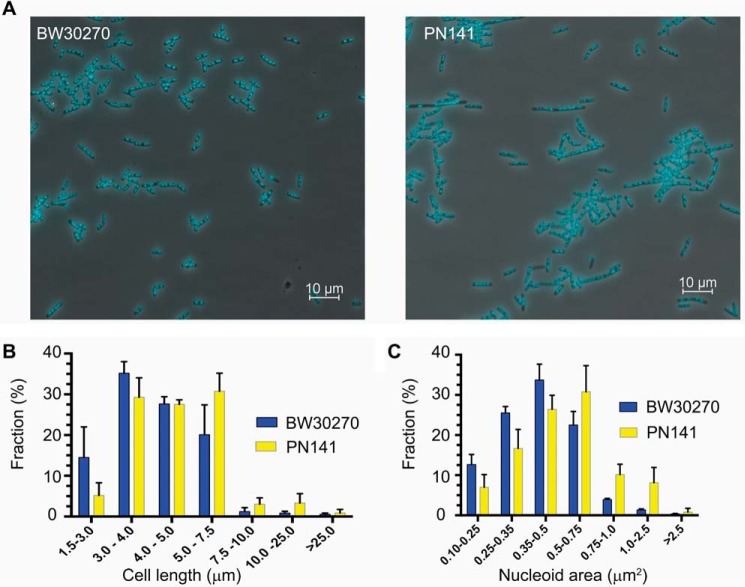
MukB is a structural maintenance of chromosome-like protein required for DNA condensation. The complete condensin is a large tripartite complex of MukB, the kleisin, MukF, and an accessory protein, MukE. As found previously, MukB DNA condensation is a stepwise process. We have defined these steps topologically. They proceed first via the formation of negative supercoils that are sequestered by the protein followed by hinge-hinge interactions between MukB dimers that stabilize topologically isolated loops in the DNA. MukB itself is sufficient to mediate both of these topological alterations; neither ATP nor MukEF is required. We show that the MukB hinge region binds DNA and that this region of the protein is involved in sequestration of supercoils. Cells carrying mutations in the MukB hinge that reduce DNA condensation in vitro exhibit nucleoid decondensation in vivo.
Keywords: DNA; DNA enzymes; DNA structure; DNA topology; chromosomes; nucleic acid enzymes; nucleic acid enzymology; nucleic acids.
© 2017 by The American Society for Biochemistry and Molecular Biology, Inc.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
Similar articles
-
The MukB-topoisomerase IV interaction is required for proper chromosome compaction.J Biol Chem. 2017 Oct 13;292(41):16921-16932. doi: 10.1074/jbc.M117.803346. Epub 2017 Aug 25. J Biol Chem. 2017. PMID: 28842485 Free PMC article.
-
MukB-mediated Catenation of DNA Is ATP and MukEF Independent.J Biol Chem. 2016 Nov 11;291(46):23999-24008. doi: 10.1074/jbc.M116.749994. Epub 2016 Oct 3. J Biol Chem. 2016. PMID: 27697840 Free PMC article.
-
The role of MukE in assembling a functional MukBEF complex.J Mol Biol. 2011 Sep 30;412(4):578-90. doi: 10.1016/j.jmb.2011.08.009. Epub 2011 Aug 10. J Mol Biol. 2011. PMID: 21855551 Free PMC article.
-
Partitioning of the Escherichia coli chromosome: superhelicity and condensation.Biochimie. 2001 Jan;83(1):41-8. doi: 10.1016/s0300-9084(00)01204-9. Biochimie. 2001. PMID: 11254973 Review.
-
New genes with old modus operandi. The connection between supercoiling and partitioning of DNA in Escherichia coli.EMBO Rep. 2000 Oct;1(4):323-7. doi: 10.1093/embo-reports/kvd077. EMBO Rep. 2000. PMID: 11269497 Free PMC article. Review.
Cited by
-
The MukB-topoisomerase IV interaction mutually suppresses their catalytic activities.Nucleic Acids Res. 2022 Mar 21;50(5):2621-2634. doi: 10.1093/nar/gkab1027. Nucleic Acids Res. 2022. PMID: 34747485 Free PMC article.
-
MukB ATPases are regulated independently by the N- and C-terminal domains of MukF kleisin.Elife. 2018 Jan 11;7:e31522. doi: 10.7554/eLife.31522. Elife. 2018. PMID: 29323635 Free PMC article.
-
Self-organised segregation of bacterial chromosomal origins.Elife. 2019 Aug 9;8:e46564. doi: 10.7554/eLife.46564. Elife. 2019. PMID: 31397672 Free PMC article.
-
Bridging-induced phase separation induced by cohesin SMC protein complexes.Sci Adv. 2021 Feb 10;7(7):eabe5905. doi: 10.1126/sciadv.abe5905. Print 2021 Feb. Sci Adv. 2021. PMID: 33568486 Free PMC article.
-
Dissecting DNA Compaction by the Bacterial Condensin MukB.Methods Mol Biol. 2019;2004:169-180. doi: 10.1007/978-1-4939-9520-2_13. Methods Mol Biol. 2019. PMID: 31147917 Free PMC article.
References
-
- Uhlmann F. (2016) SMC complexes: from DNA to chromosomes. Nat. Rev. Mol. Cell Biol. 17, 399–412 - PubMed
-
- Yamanaka K., Ogura T., Niki H., and Hiraga S. (1996) Identification of two new genes, mukE and mukF, involved in chromosome partitioning in Escherichia coli. Mol. Gen. Genet. 250, 241–251 - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
