

Distinct Mechanism Evolved for Mycobacterial RNA Polymerase and Topoisomerase I Protein-Protein Interaction
- PMID: 28843989
- PMCID: PMC5610943
- DOI: 10.1016/j.jmb.2017.08.011
Distinct Mechanism Evolved for Mycobacterial RNA Polymerase and Topoisomerase I Protein-Protein Interaction
Abstract
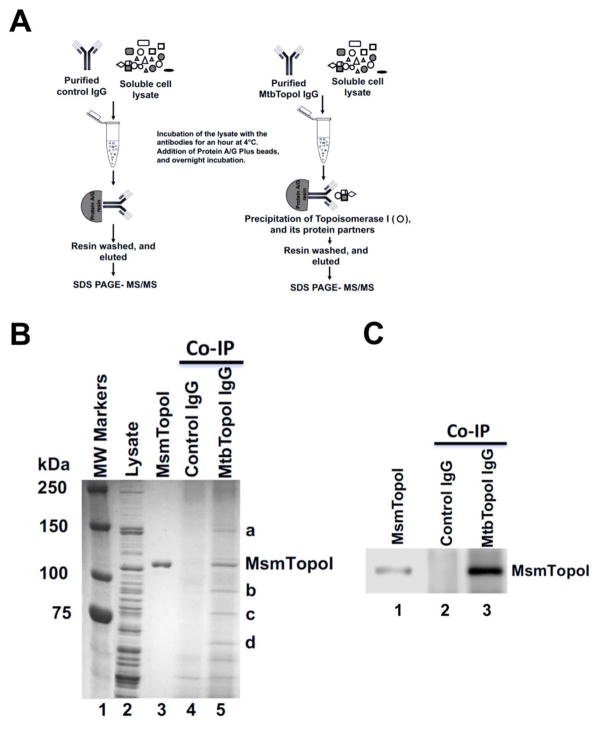
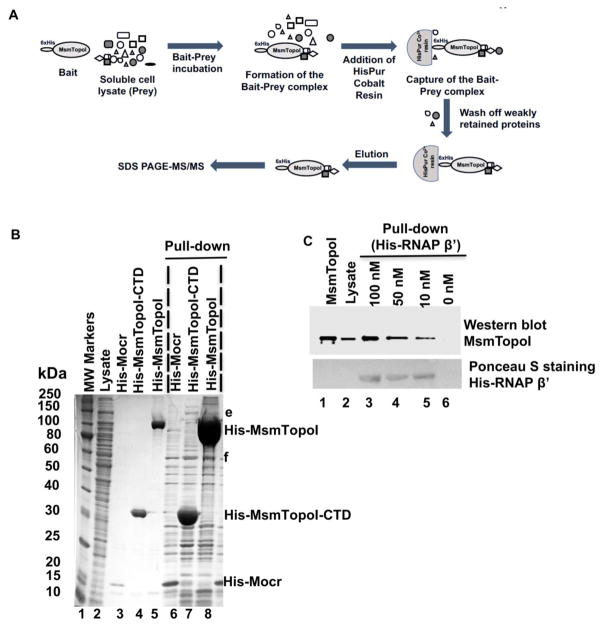
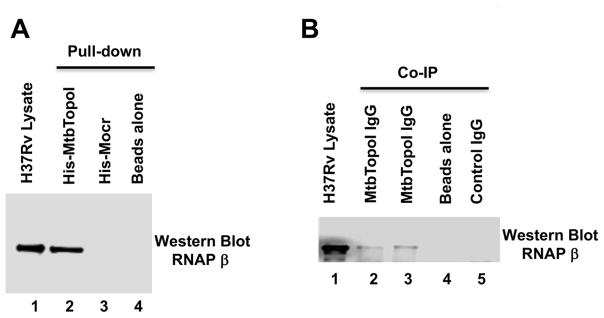
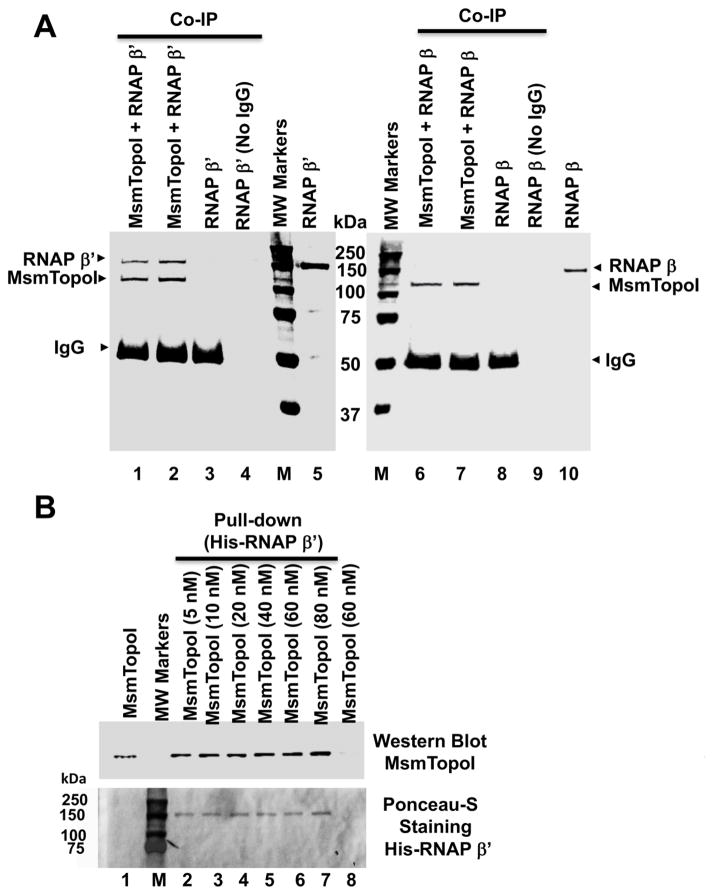
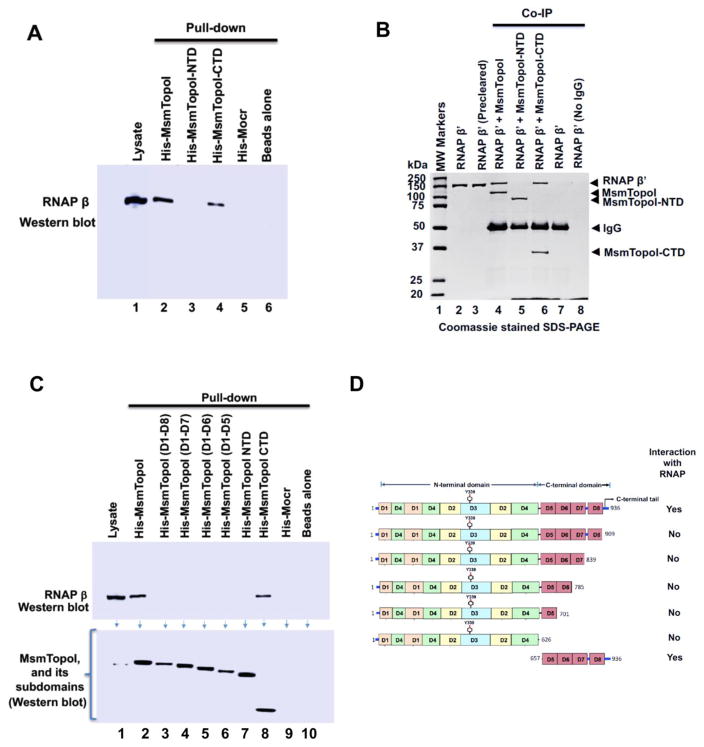
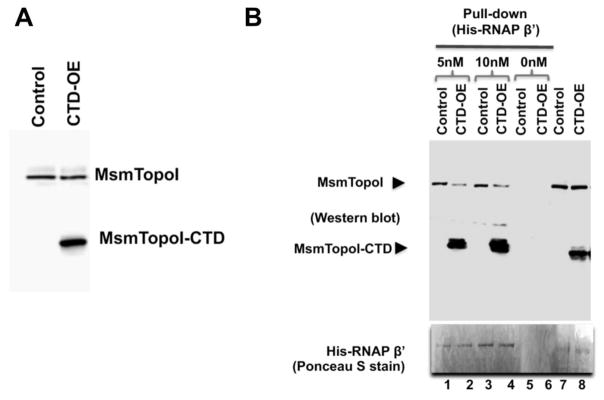
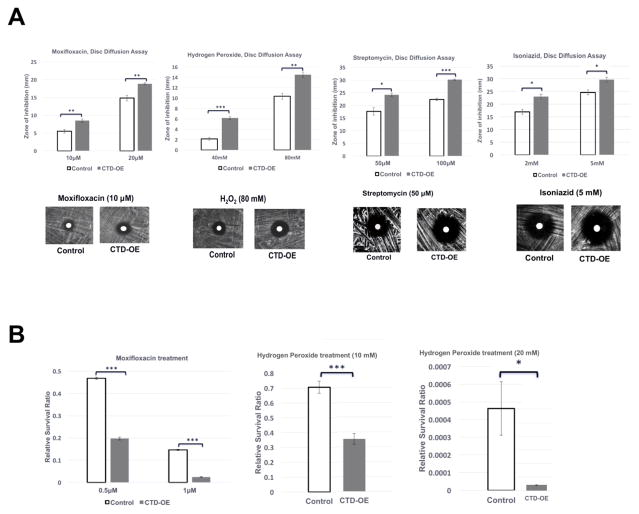
We report here a distinct mechanism of interaction between topoisomerase I and RNA polymerase in Mycobacterium tuberculosis and Mycobacterium smegmatis that has evolved independently from the previously characterized interaction between bacterial topoisomerase I and RNA polymerase. Bacterial DNA topoisomerase I is responsible for preventing the hyper-negative supercoiling of genomic DNA. The association of topoisomerase I with RNA polymerase during transcription elongation could efficiently relieve transcription-driven negative supercoiling. Our results demonstrate a direct physical interaction between the C-terminal domains of topoisomerase I (TopoI-CTDs) and the β' subunit of RNA polymerase of M. smegmatis in the absence of DNA. The TopoI-CTDs in mycobacteria are evolutionarily unrelated in amino acid sequence and three-dimensional structure to the TopoI-CTD found in the majority of bacterial species outside Actinobacteria, including Escherichia coli. The functional interaction between topoisomerase I and RNA polymerase has evolved independently in mycobacteria and E. coli, with distinctively different structural elements of TopoI-CTD utilized for this protein-protein interaction. Zinc ribbon motifs in E. coli TopoI-CTD are involved in the interaction with RNA polymerase. For M. smegmatis TopoI-CTD, a 27-amino-acid tail that is rich in basic residues at the C-terminal end is responsible for the interaction with RNA polymerase. Overexpression of recombinant TopoI-CTD in M. smegmatis competed with the endogenous topoisomerase I for protein-protein interactions with RNA polymerase. The TopoI-CTD overexpression resulted in decreased survival following treatment with antibiotics and hydrogen peroxide, supporting the importance of the protein-protein interaction between topoisomerase I and RNA polymerase during stress response of mycobacteria.
Keywords: TB; antibiotic sensitivity; evolution; stress response; transcription elongation.
Copyright © 2017 Elsevier Ltd. All rights reserved.
Figures
Similar articles
-
Autoregulation of topoisomerase I expression by supercoiling sensitive transcription.Nucleic Acids Res. 2016 Feb 29;44(4):1541-52. doi: 10.1093/nar/gkv1088. Epub 2015 Oct 22. Nucleic Acids Res. 2016. PMID: 26496944 Free PMC article.
-
Antimicrobial Susceptibility and SOS-Dependent Increase in Mutation Frequency Are Impacted by Escherichia coli Topoisomerase I C-Terminal Point Mutation.Antimicrob Agents Chemother. 2015 Oct;59(10):6195-202. doi: 10.1128/AAC.00855-15. Epub 2015 Jul 27. Antimicrob Agents Chemother. 2015. PMID: 26248366 Free PMC article.
-
Direct interaction between Escherichia coli RNA polymerase and the zinc ribbon domains of DNA topoisomerase I.J Biol Chem. 2003 Aug 15;278(33):30705-10. doi: 10.1074/jbc.M303403200. Epub 2003 Jun 4. J Biol Chem. 2003. PMID: 12788950
-
Multifunctional Mycobacterial Topoisomerases with Distinctive Features.ACS Infect Dis. 2025 Feb 14;11(2):366-385. doi: 10.1021/acsinfecdis.4c00880. Epub 2025 Jan 18. ACS Infect Dis. 2025. PMID: 39825760 Review.
-
[DNA supercoiling and topoisomerases in Escherichia coli].Rev Latinoam Microbiol. 1995 Jul-Sep;37(3):291-304. Rev Latinoam Microbiol. 1995. PMID: 8850348 Review. Spanish.
Cited by
-
Genome-wide proximity between RNA polymerase and DNA topoisomerase I supports transcription in Streptococcus pneumoniae.PLoS Genet. 2021 Apr 30;17(4):e1009542. doi: 10.1371/journal.pgen.1009542. eCollection 2021 Apr. PLoS Genet. 2021. PMID: 33930020 Free PMC article.
-
Interaction between transcribing RNA polymerase and topoisomerase I prevents R-loop formation in E. coli.Nat Commun. 2022 Aug 4;13(1):4524. doi: 10.1038/s41467-022-32106-5. Nat Commun. 2022. PMID: 35927234 Free PMC article.
-
Mechanism of Type IA Topoisomerases.Molecules. 2020 Oct 17;25(20):4769. doi: 10.3390/molecules25204769. Molecules. 2020. PMID: 33080770 Free PMC article. Review.
-
Microbial Type IA Topoisomerase C-Terminal Domain Sequence Motifs, Distribution and Combination.Int J Mol Sci. 2022 Aug 5;23(15):8709. doi: 10.3390/ijms23158709. Int J Mol Sci. 2022. PMID: 35955842 Free PMC article.
-
Supercoiling, R-loops, Replication and the Functions of Bacterial Type 1A Topoisomerases.Genes (Basel). 2020 Feb 27;11(3):249. doi: 10.3390/genes11030249. Genes (Basel). 2020. PMID: 32120891 Free PMC article. Review.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
