

Transposable Element Domestication As an Adaptation to Evolutionary Conflicts
- PMID: 28844698
- PMCID: PMC5659911
- DOI: 10.1016/j.tig.2017.07.011
Transposable Element Domestication As an Adaptation to Evolutionary Conflicts
Abstract
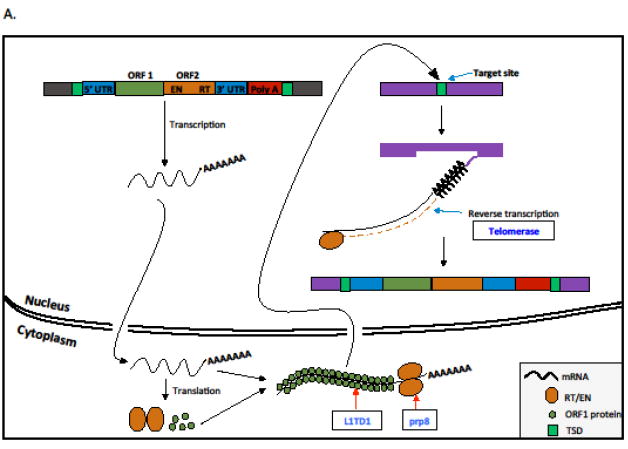
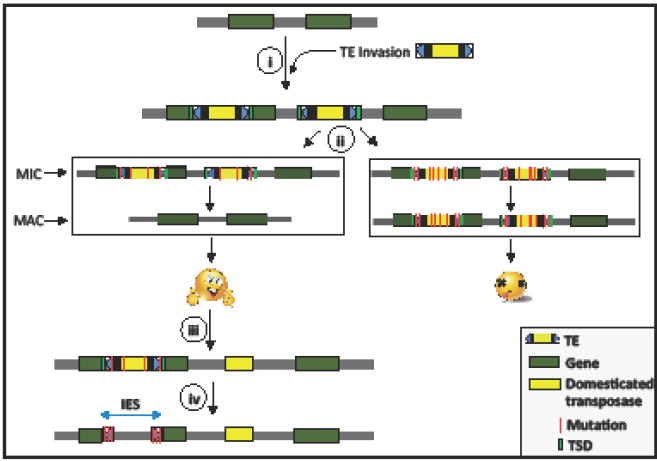
Transposable elements (TEs) are selfish genetic units that typically encode proteins that enable their proliferation in the genome and spread across individual hosts. Here we review a growing number of studies that suggest that TE proteins have often been co-opted or 'domesticated' by their host as adaptations to a variety of evolutionary conflicts. In particular, TE-derived proteins have been recurrently repurposed as part of defense systems that protect prokaryotes and eukaryotes against the proliferation of infectious or invasive agents, including viruses and TEs themselves. We argue that the domestication of TE proteins may often be the only evolutionary path toward the mitigation of the cost incurred by their own selfish activities.
Keywords: adaptation; evolutionary conflicts; transposable element protein domestication.
Copyright © 2017 Elsevier Ltd. All rights reserved.
Figures
References
-
- Curcio MJ, Derbyshire KM. The outs and ins of transposition: from mu to kangaroo. Nat Rev Mol Cell Biol. 2003;4(11):865–77. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
