

Nuclear DNA Replication in Trypanosomatids: There Are No Easy Methods for Solving Difficult Problems
- PMID: 28844718
- PMCID: PMC5662062
- DOI: 10.1016/j.pt.2017.08.002
Nuclear DNA Replication in Trypanosomatids: There Are No Easy Methods for Solving Difficult Problems
Abstract
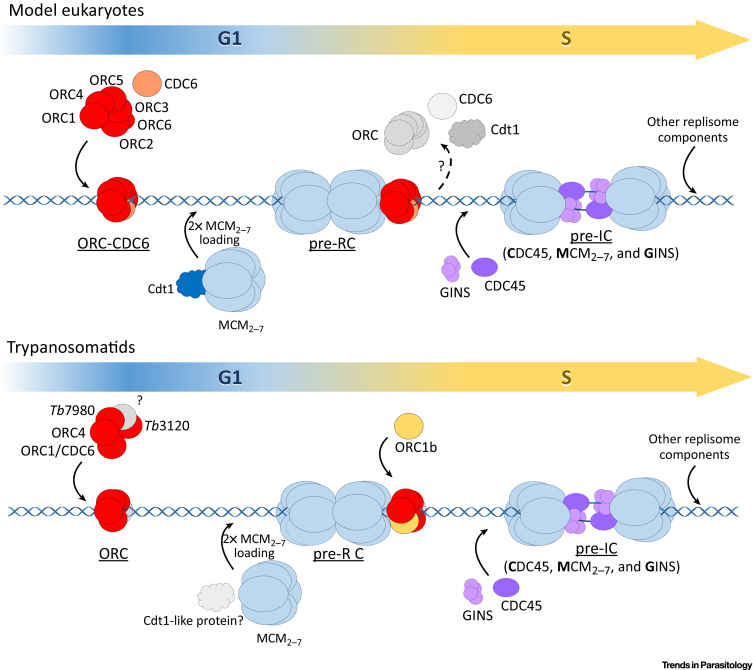
In trypanosomatids, etiological agents of devastating diseases, replication is robust and finely controlled to maintain genome stability and function in stressful environments. However, these parasites encode several replication protein components and complexes that show potentially variant composition compared with model eukaryotes. This review focuses on the advances made in recent years regarding the differences and peculiarities of the replication machinery in trypanosomatids, including how such divergence might affect DNA replication dynamics and the replication stress response. Comparing the DNA replication machinery and processes of parasites and their hosts may provide a foundation for the identification of targets that can be used in the development of chemotherapies to assist in the eradication of diseases caused by these pathogens.
Keywords: DNA replication; origin recognition complex; replication fork; replication origins; replication stress; trypanosomatid emergence.
Copyright © 2017 The Authors. Published by Elsevier Ltd.. All rights reserved.
Figures





Comment in
-
Maria Carolina Elias: Parasites with Dynamic Genomes.Trends Parasitol. 2017 Nov;33(11):827-828. doi: 10.1016/j.pt.2017.08.008. Epub 2017 Sep 11. Trends Parasitol. 2017. PMID: 28919219 No abstract available.
References
-
- da Silva M.S. Differences in the detection of BrdU/EdU incorporation assays alter the calculation for G1, S, and G2 phases of the cell cycle in trypanosomatids. J. Eukaryot. Microbiol. 2017 Published online March 4, 2017. - PubMed
-
- El-Sayed N.M. Comparative genomics of trypanosomatid parasitic protozoa. Science. 2005;309:404–409. - PubMed
-
- Simpson A. Early evolution within kinetoplastids (Euglenozoa), and the late emergence of trypanosomatids. Protist. 2004;155:407–422. - PubMed
-
- Flegontov P. Paratrypanosoma is a novel early-branching trypanosomatid. Curr. Biol. 2013;23:1787–1793. - PubMed
-
- Simpson A.G.B. The evolution and diversity of kinetoplastid flagellates. Trends Parasitol. 2006;22:168–174. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
