

Monitoring and Measuring Autophagy
- PMID: 28846632
- PMCID: PMC5618514
- DOI: 10.3390/ijms18091865
Monitoring and Measuring Autophagy
Abstract
Autophagy is a cytoplasmic degradation system, which is important for starvation adaptation and cellular quality control. Recent advances in understanding autophagy highlight its importance under physiological and pathological conditions. However, methods for monitoring autophagic activity are complicated and the results are sometimes misinterpreted. Here, we review the methods used to identify autophagic structures, and to measure autophagic flux in cultured cells and animals. We will also describe the existing autophagy reporter mice that are useful for autophagy studies and drug testing. Lastly, we will consider the attempts to monitor autophagy in samples derived from humans.
Keywords: Keima; LC3; autophagic flux; autophagosome; isolation membrane; p62/SQSTM1.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures
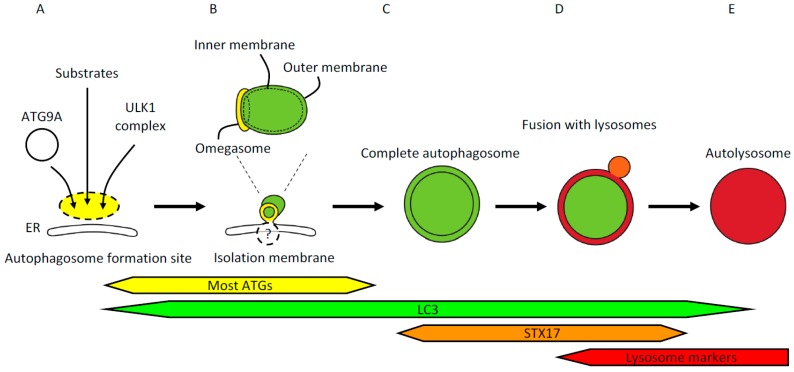
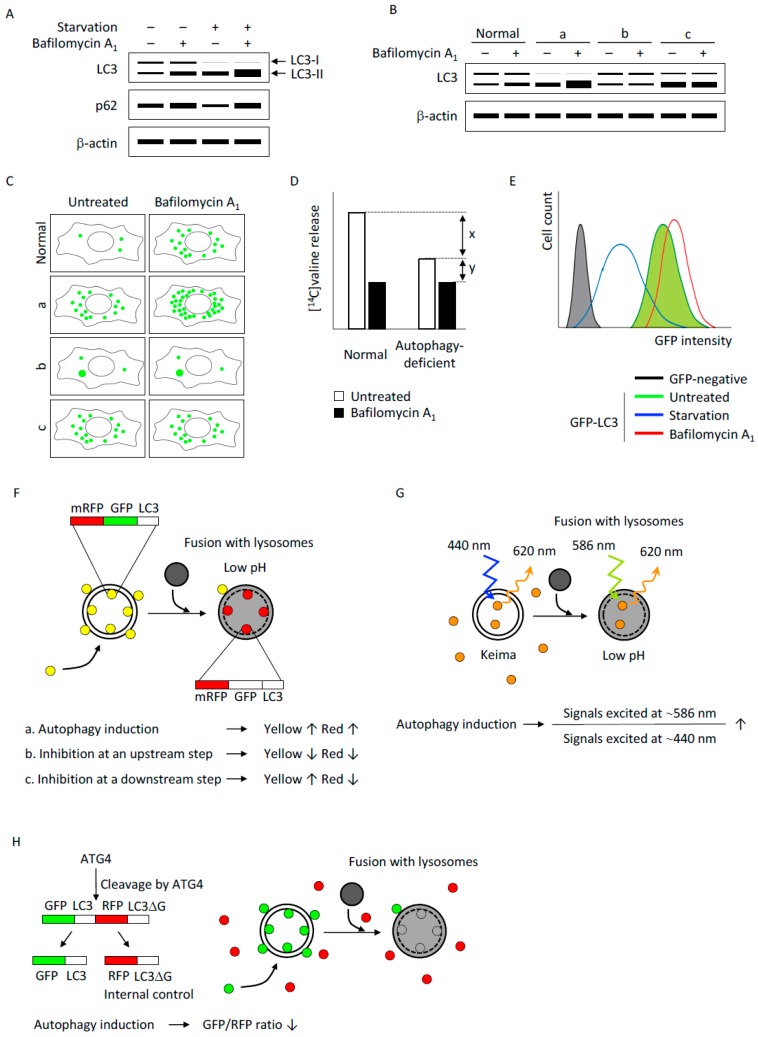
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
