

CTCF counter-regulates cardiomyocyte development and maturation programs in the embryonic heart
- PMID: 28846746
- PMCID: PMC5591014
- DOI: 10.1371/journal.pgen.1006985
CTCF counter-regulates cardiomyocyte development and maturation programs in the embryonic heart
Abstract
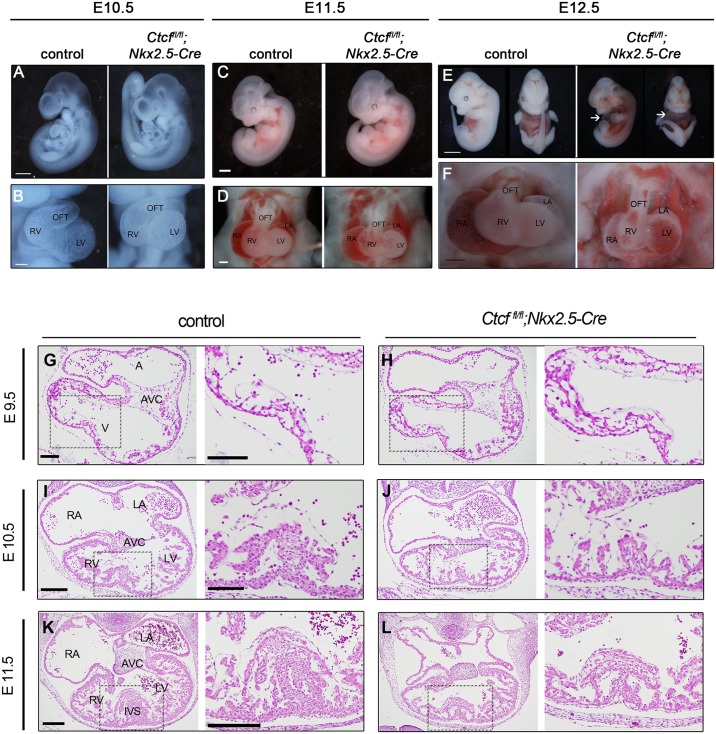
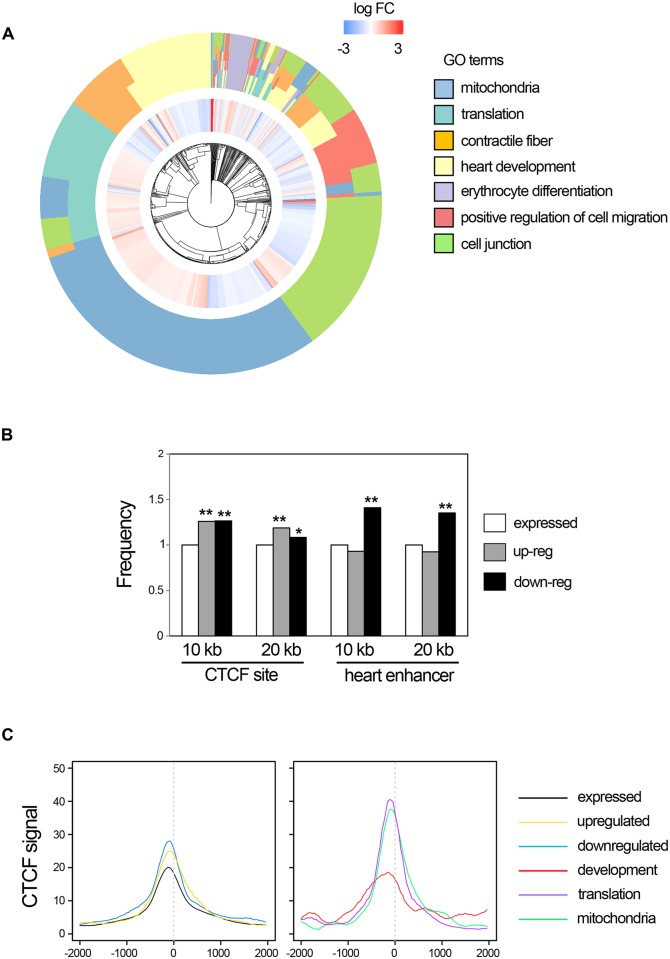
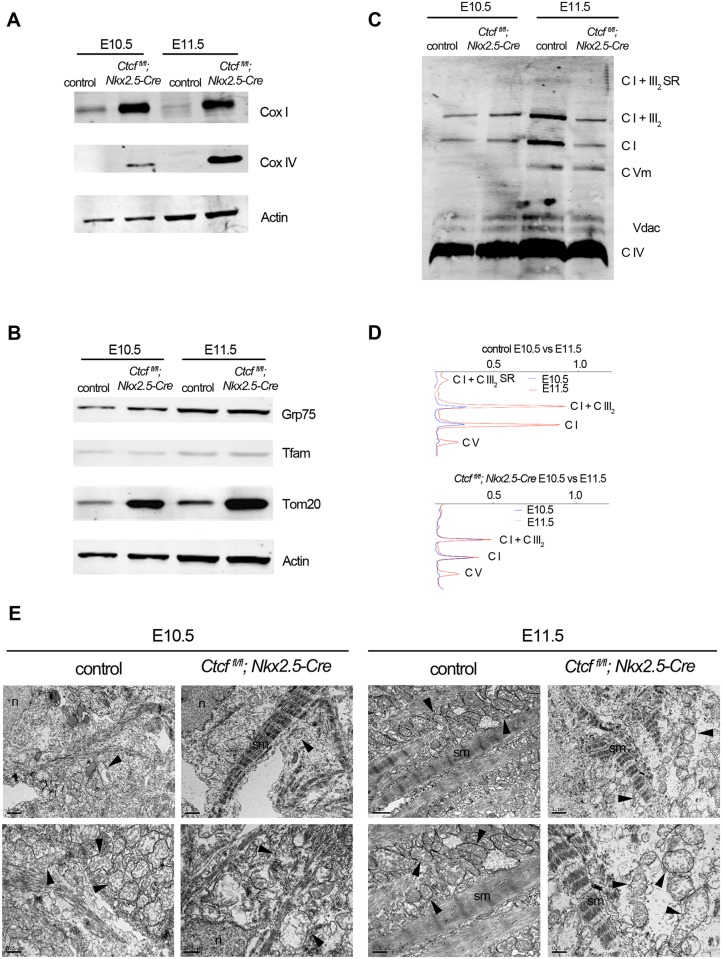
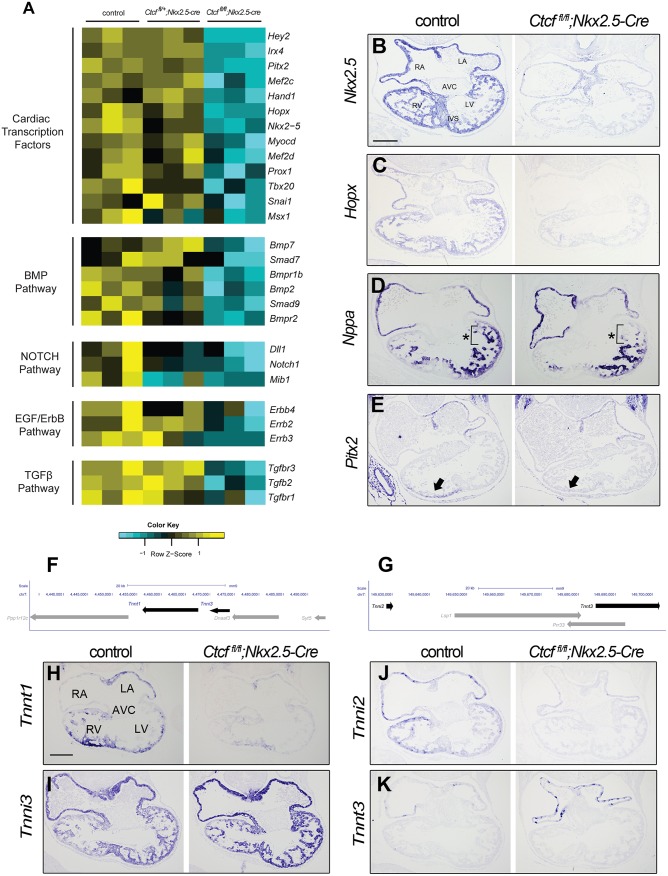
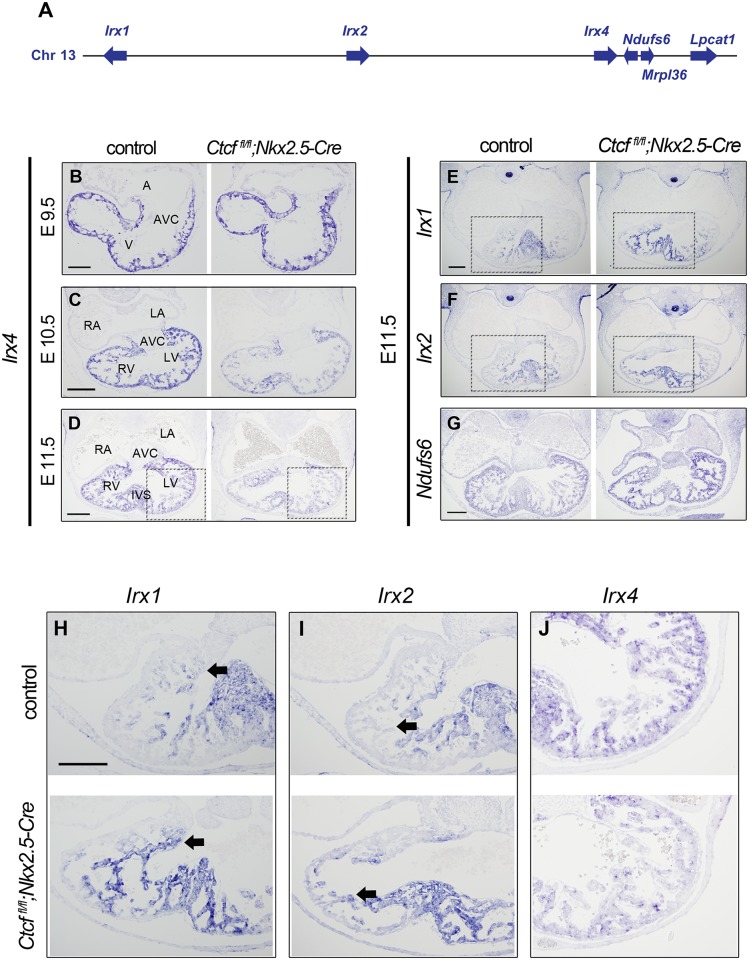
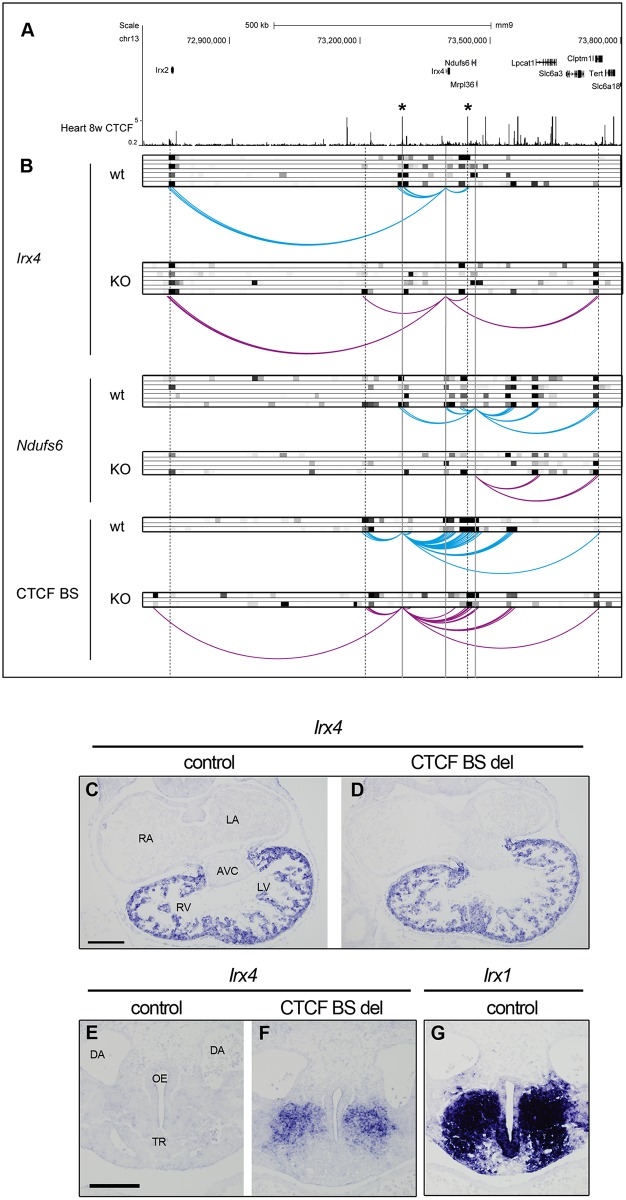
Cardiac progenitors are specified early in development and progressively differentiate and mature into fully functional cardiomyocytes. This process is controlled by an extensively studied transcriptional program. However, the regulatory events coordinating the progression of such program from development to maturation are largely unknown. Here, we show that the genome organizer CTCF is essential for cardiogenesis and that it mediates genomic interactions to coordinate cardiomyocyte differentiation and maturation in the developing heart. Inactivation of Ctcf in cardiac progenitor cells and their derivatives in vivo during development caused severe cardiac defects and death at embryonic day 12.5. Genome wide expression analysis in Ctcf mutant hearts revealed that genes controlling mitochondrial function and protein production, required for cardiomyocyte maturation, were upregulated. However, mitochondria from mutant cardiomyocytes do not mature properly. In contrast, multiple development regulatory genes near predicted heart enhancers, including genes in the IrxA cluster, were downregulated in Ctcf mutants, suggesting that CTCF promotes cardiomyocyte differentiation by facilitating enhancer-promoter interactions. Accordingly, loss of CTCF disrupts gene expression and chromatin interactions as shown by chromatin conformation capture followed by deep sequencing. Furthermore, CRISPR-mediated deletion of an intergenic CTCF site within the IrxA cluster alters gene expression in the developing heart. Thus, CTCF mediates local regulatory interactions to coordinate transcriptional programs controlling transitions in morphology and function during heart development.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Olson EN. Gene regulatory networks in the evolution and development of the heart. Science. 2006;313(5795):1922–7. doi: 10.1126/science.1132292 - DOI - PMC - PubMed
-
- Hirschy A, Schatzmann F, Ehler E, Perriard JC. Establishment of cardiac cytoarchitecture in the developing mouse heart. Dev Biol. 2006;289(2):430–41. doi: 10.1016/j.ydbio.2005.10.046 - DOI - PubMed
-
- Hom JR, Quintanilla RA, Hoffman DL, de Mesy Bentley KL, Molkentin JD, Sheu SS, et al. The permeability transition pore controls cardiac mitochondrial maturation and myocyte differentiation. Dev Cell. 2011;21(3):469–78. doi: 10.1016/j.devcel.2011.08.008 - DOI - PMC - PubMed
-
- Beutner G, Eliseev RA, Porter GA Jr. Initiation of electron transport chain activity in the embryonic heart coincides with the activation of mitochondrial complex 1 and the formation of supercomplexes. PLoS One. 2014;9(11):e113330 doi: 10.1371/journal.pone.0113330 - DOI - PMC - PubMed
-
- Soonpaa MH, Kim KK, Pajak L, Franklin M, Field LJ. Cardiomyocyte DNA synthesis and binucleation during murine development. Am J Physiol. 1996;271(5 Pt 2):H2183–9. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
