

The Tumor Necrosis Factor Family: Family Conventions and Private Idiosyncrasies
- PMID: 28847899
- PMCID: PMC6169814
- DOI: 10.1101/cshperspect.a028431
The Tumor Necrosis Factor Family: Family Conventions and Private Idiosyncrasies
Abstract
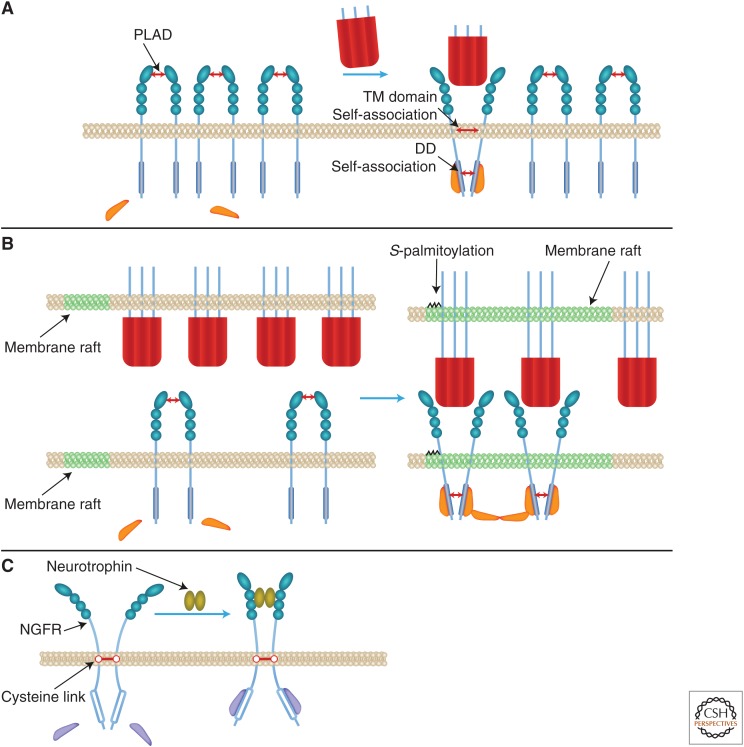
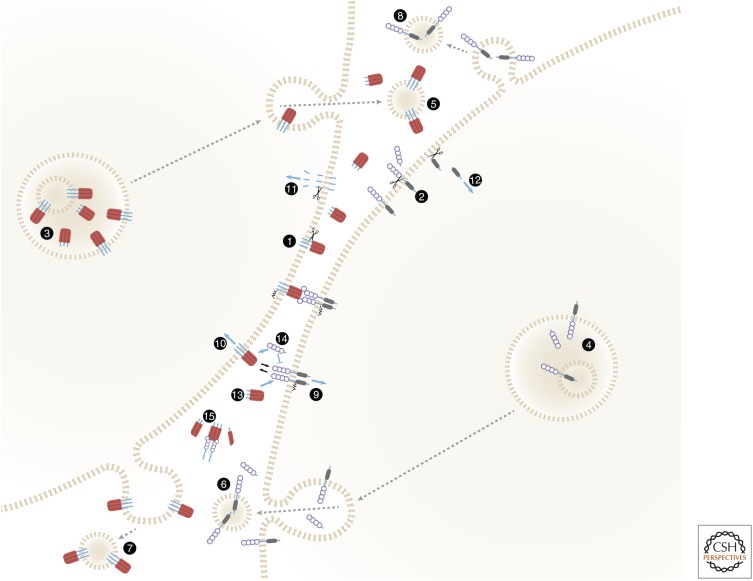
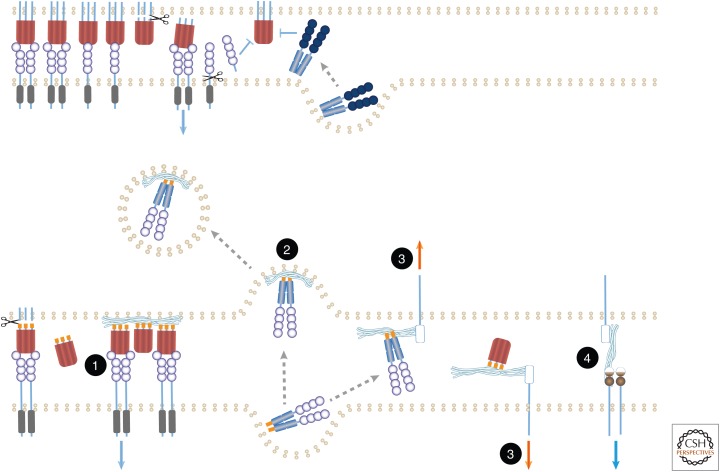
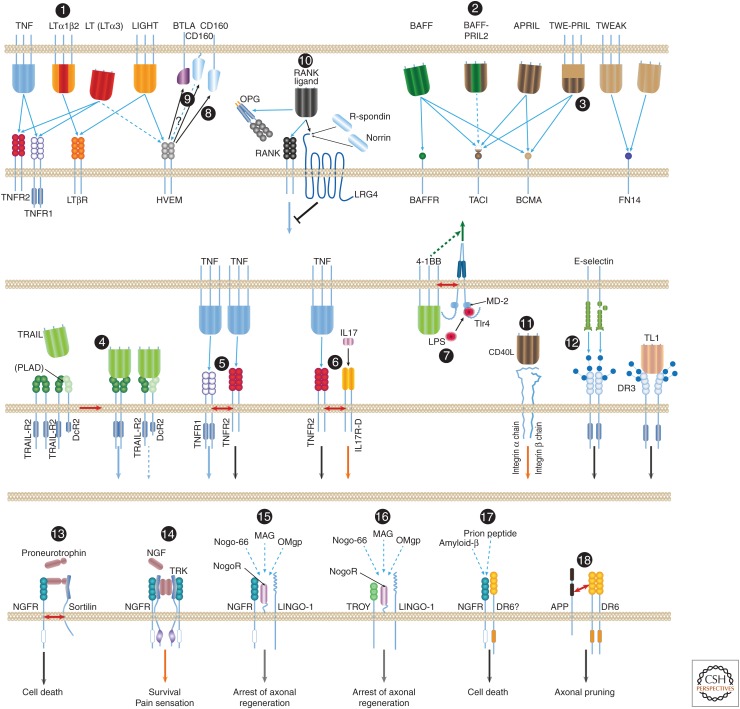
The tumor necrosis factor (TNF) cytokine family and the TNF/nerve growth factor (NGF) family of their cognate receptors together control numerous immune functions, as well as tissue-homeostatic and embryonic-development processes. These diverse functions are dictated by both shared and distinct features of family members, and by interactions of some members with nonfamily ligands and coreceptors. The spectra of their activities are further expanded by the occurrence of the ligands and receptors in both membrane-anchored and soluble forms, by "re-anchoring" of soluble forms to extracellular matrix components, and by signaling initiation via intracellular domains (IDs) of both receptors and ligands. Much has been learned about shared features of the receptors as well as of the ligands; however, we still have only limited knowledge of the mechanistic basis for their functional heterogeneity and for the differences between their functions and those of similarly acting cytokines of other families.
Copyright © 2018 Cold Spring Harbor Laboratory Press; all rights reserved.
Figures
References
-
- Adam Klages S, Adam D, Wiegmann K, Struve S, Kolanus W, Schneider Mergener J, Kronke M. 1996. FAN, a novel WD-repeat protein, couples the p55 TNF-receptor to neutral sphingomyelinase. Cell 86: 937–947. - PubMed
-
- Akassoglou K, Bauer J, Kassiotis G, Lassmann H, Kollias G, Probert L. 1999. Transgenic models of TNF induced demyelination. Adv Exp Med Biol 468: 245–259. - PubMed
-
- Albanese J, Meterissian S, Kontogiannea M, Dubreuil C, Hand A, Sorba S, Dainiak N. 1998. Biologically active Fas antigen and its cognate ligand are expressed on plasma membrane-derived extracellular vesicles. Blood 91: 3862–3874. - PubMed
-
- Alon R, Cahalon L, Hershkoviz R, Elbaz D, Reizis B, Wallach D, Akiyama SK, Yamada KM, Lider O. 1994. TNF-α binds to the N-terminal domain of fibronectin and augments the β1-integrin-mediated adhesion of CD4+ T lymphocytes to the glycoprotein. J Immunol 152: 1304–1313. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases