

Biochemical Mechanisms and Microorganisms Involved in Anaerobic Testosterone Metabolism in Estuarine Sediments
- PMID: 28848528
- PMCID: PMC5554518
- DOI: 10.3389/fmicb.2017.01520
Biochemical Mechanisms and Microorganisms Involved in Anaerobic Testosterone Metabolism in Estuarine Sediments
Abstract
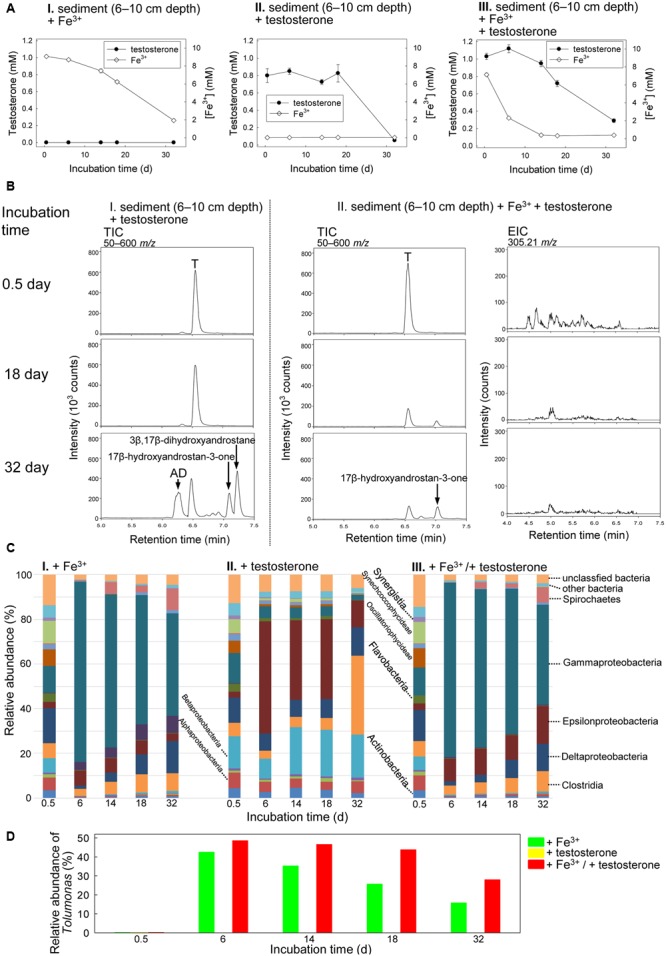
Current knowledge on the biochemical mechanisms underlying microbial steroid metabolism in anaerobic ecosystems is extremely limited. Sulfate, nitrate, and iron [Fe (III)] are common electron acceptors for anaerobes in estuarine sediments. Here, we investigated anaerobic testosterone metabolism in anaerobic sediments collected from the estuary of Tamsui River, Taiwan. The anaerobic sediment samples were spiked with testosterone (1 mM) and individual electron acceptors (10 mM), including nitrate, Fe3+, and sulfate. The analysis of androgen metabolites indicated that testosterone biodegradation under denitrifying conditions proceeds through the 2,3-seco pathway, whereas testosterone biodegradation under iron-reducing conditions may proceed through an unidentified alternative pathway. Metagenomic analysis and PCR-based functional assays suggested that Thauera spp. were the major testosterone degraders in estuarine sediment samples incubated with testosterone and nitrate. Thauera sp. strain GDN1, a testosterone-degrading betaproteobacterium, was isolated from the denitrifying sediment sample. This strain tolerates a broad range of salinity (0-30 ppt). Although testosterone biodegradation did not occur under sulfate-reducing conditions, we observed the anaerobic biotransformation of testosterone to estrogens in some testosterone-spiked sediment samples. This is unprecedented since biotransformation of androgens to estrogens is known to occur only under oxic conditions. Our metagenomic analysis suggested that Clostridium spp. might play a role in this anaerobic biotransformation. These results expand our understanding of microbial metabolism of steroids under strictly anoxic conditions.
Keywords: Illumina MiSeq; Thauera; androgen; biodegradation; estrogen; estuary; sediment; testosterone.
Figures





References
-
- Boyle E. A., Edmond J. M., Sholkovitz E. R. (1977). The mechanism of iron removal in estuaries. Geochim. Cosmochim. Acta 41 1313–1324. 10.1021/acs.est.5b03419 - DOI
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous