

Swiss Army Pathogen: The Salmonella Entry Toolkit
- PMID: 28848711
- PMCID: PMC5552672
- DOI: 10.3389/fcimb.2017.00348
Swiss Army Pathogen: The Salmonella Entry Toolkit
Abstract
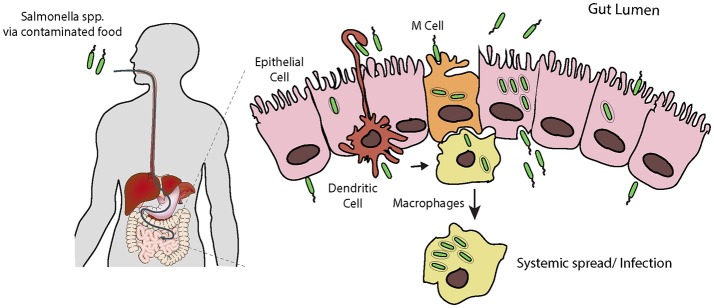
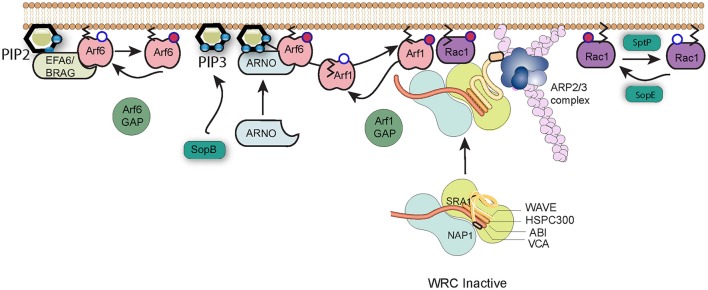
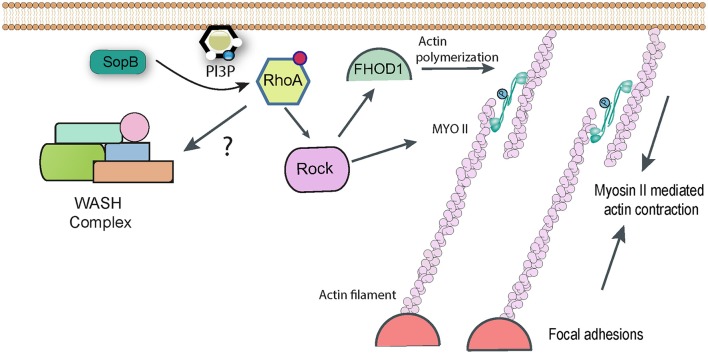
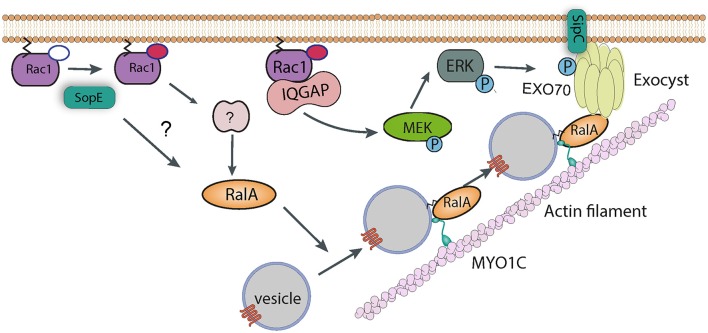
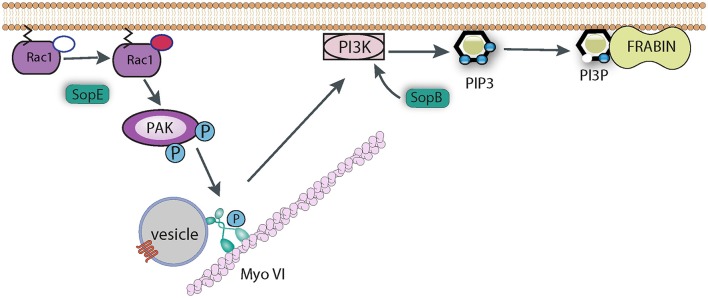
Salmonella causes disease in humans and animals ranging from mild self-limiting gastroenteritis to potentially life-threatening typhoid fever. Salmonellosis remains a considerable cause of morbidity and mortality globally, and hence imposes a huge socio-economic burden worldwide. A key property of all pathogenic Salmonella strains is the ability to invade non-phagocytic host cells. The major determinant of this invasiveness is a Type 3 Secretion System (T3SS), a molecular syringe that injects virulence effector proteins directly into target host cells. These effectors cooperatively manipulate multiple host cell signaling pathways to drive pathogen internalization. Salmonella does not only rely on these injected effectors, but also uses several other T3SS-independent mechanisms to gain entry into host cells. This review summarizes our current understanding of the methods used by Salmonella for cell invasion, with a focus on the host signaling networks that must be coordinately exploited for the pathogen to achieve its goal.
Keywords: SPI1-independent entry; Salmonella invasion; Salmonella pathogenicity islands; T3SS effectors; actin cytoskeleton; membrane ruffling.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical