

RNA-virus proteases counteracting host innate immunity
- PMID: 28850669
- PMCID: PMC7163997
- DOI: 10.1002/1873-3468.12827
RNA-virus proteases counteracting host innate immunity
Abstract
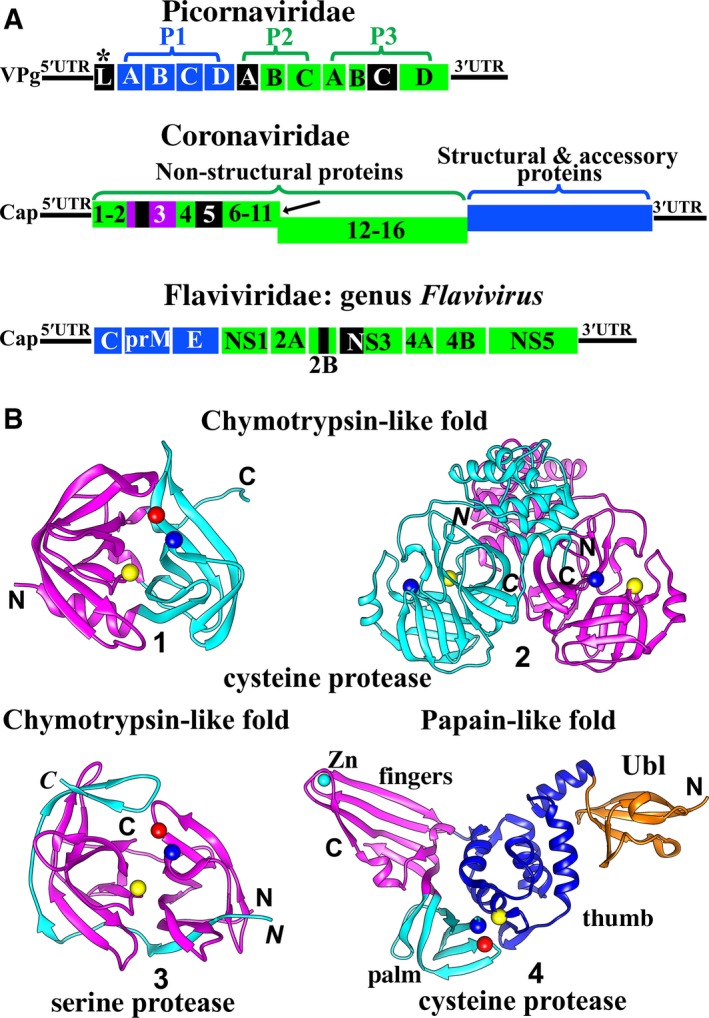
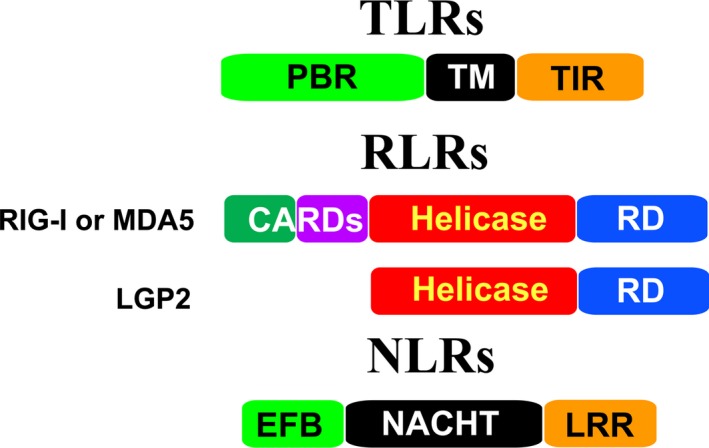
Virus invasion triggers host immune responses, in particular, innate immune responses. Pathogen-associated molecular patterns of viruses (such as dsRNA, ssRNA, or viral proteins) released during virus replication are detected by the corresponding pattern-recognition receptors of the host, and innate immune responses are induced. Through production of type-I and type-III interferons as well as various other cytokines, the host innate immune system forms the frontline to protect host cells and inhibit virus infection. Not surprisingly, viruses have evolved diverse strategies to counter this antiviral system. In this review, we discuss the multiple strategies used by proteases of positive-sense single-stranded RNA viruses of the families Picornaviridae, Coronaviridae, and Flaviviridae, when counteracting host innate immune responses.
Keywords: cleavage of host proteins; innate immunity; viral protease.
© 2017 Federation of European Biochemical Societies.
Figures

References
-
- Adams MJ, Lefkowitz EJ, King AMQ, Harrach B, Harrison RL, Knowles NJ, Kropinski AM, Krupovic M, Kuhn JH, Mushegian AR et al (2017) Changes to taxonomy and the International Code of Virus Classification and Nomenclature ratified by the International Committee on Taxonomy of Viruses. Arch Virol 162, 2505–2538. - PubMed
-
- Norder H, De Palma AM, Selisko B, Costenaro L, Papageorgiou N, Arnan C, Coutard B, Lantez V, De Lamballerie X, Baronti C et al (2011) Picornavirus non‐structural proteins as targets for new anti‐virals with broad activity. Antiviral Res 89, 204–218. - PubMed
-
- Seipelt J, Guarné A, Bergmann E, James M, Sommergruber W, Fita I and Skern T (1999) The structures of picornaviral proteinases. Virus Res 62, 159–168. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
