

Expression profiling of genes regulated by sphingosine kinase1 signaling in a murine model of hyperoxia induced neonatal bronchopulmonary dysplasia
- PMID: 28851267
- PMCID: PMC5576338
- DOI: 10.1186/s12864-017-4048-0
Expression profiling of genes regulated by sphingosine kinase1 signaling in a murine model of hyperoxia induced neonatal bronchopulmonary dysplasia
Abstract
Background: Sphingosine- 1-Phosphate (S1P) is a bioactive lipid and an intracellular as well as an extracellular signaling molecule. S1P ligand specifically binds to five related cell surface G-protein-coupled receptors (S1P1-5). S1P levels are tightly regulated by its synthesis catalyzed by sphingosine kinases (SphKs) 1 & 2 and catabolism by S1P phosphatases, lipid phosphate phosphatases and S1P lyase. We previously reported that knock down of SphK1 (Sphk1 -/- ) in a neonatal mouse BPD model conferred significant protection against hyperoxia induced lung injury. To better understand the underlying molecular mechanisms, genome-wide gene expression profiling was performed on mouse lung tissue using Affymetrix MoGene 2.0 array.
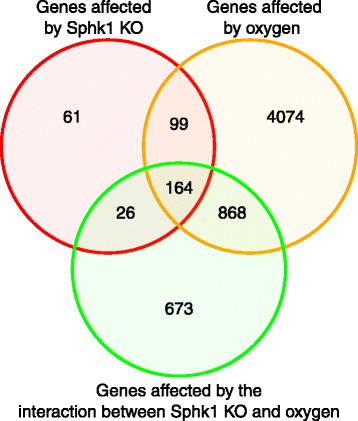
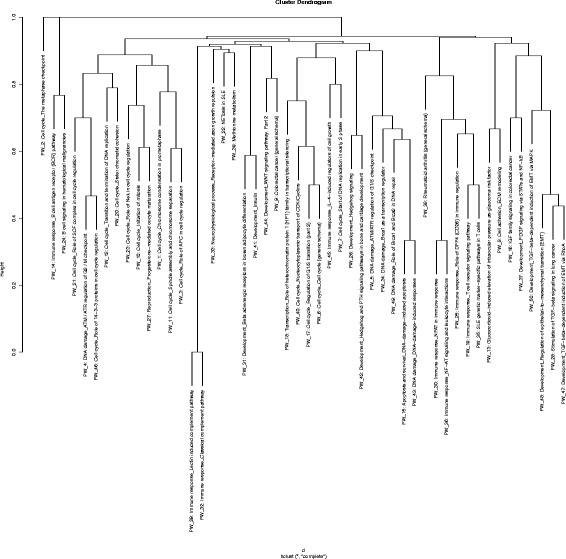
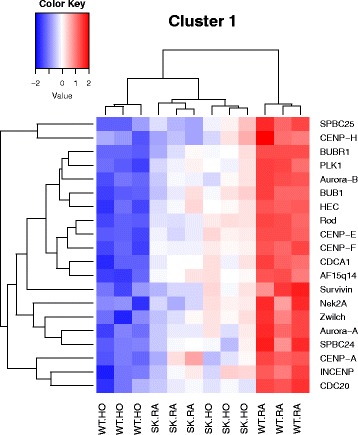
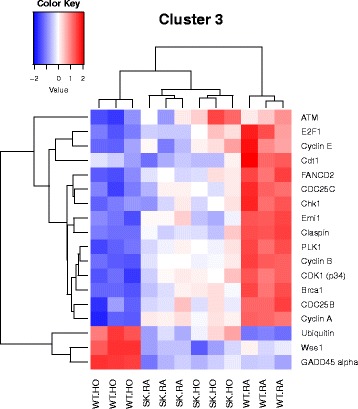
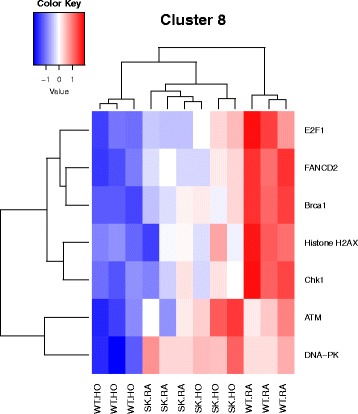
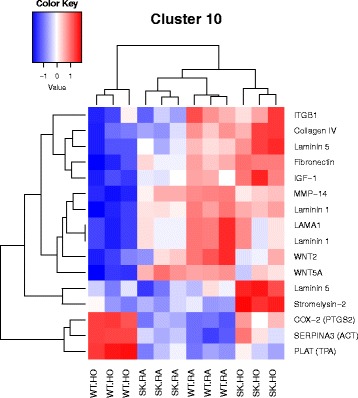
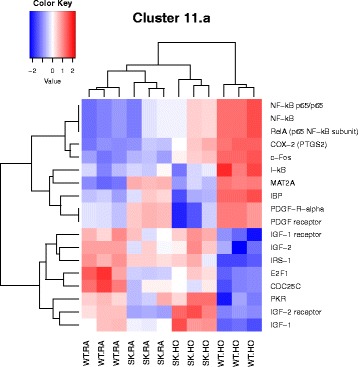
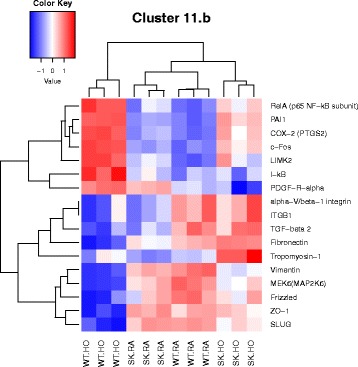
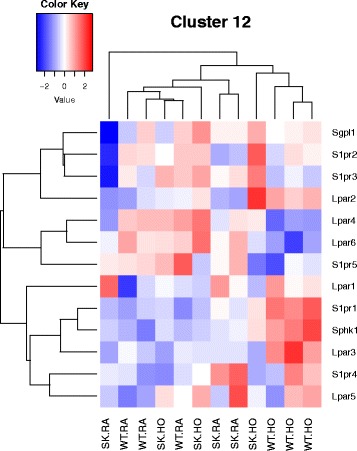
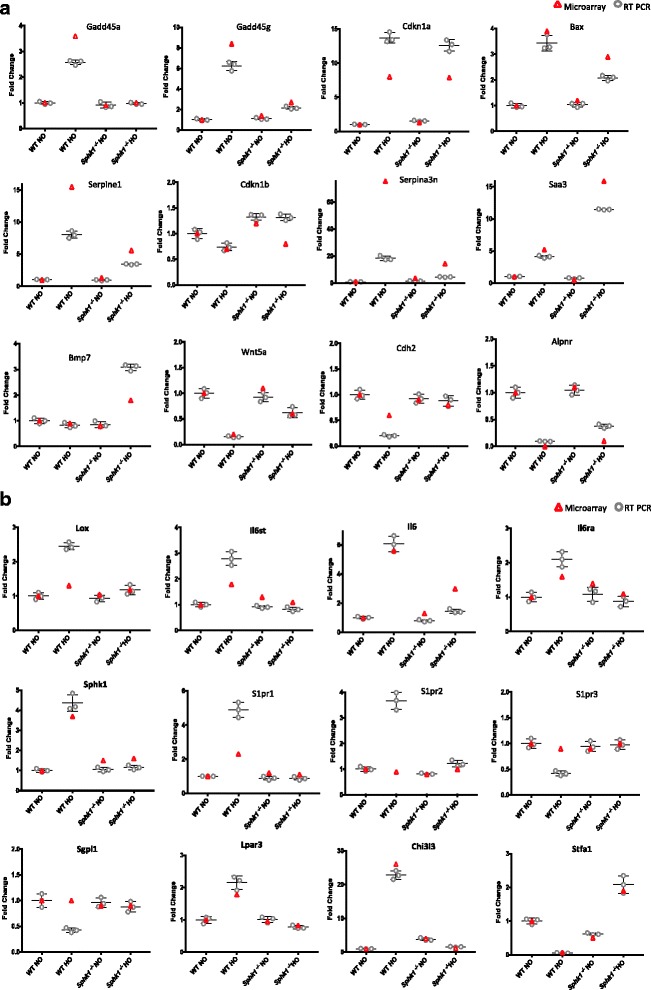
Results: Two-way ANOVA analysis was performed and differentially expressed genes under hyperoxia were identified using Sphk1 -/- mice and their wild type (WT) equivalents. Pathway (PW) enrichment analyses identified several signaling pathways that are likely to play a key role in hyperoxia induced lung injury in the neonates. These included signaling pathways that were anticipated such as those involved in lipid signaling, cell cycle regulation, DNA damage/apoptosis, inflammation/immune response, and cell adhesion/extracellular matrix (ECM) remodeling. We noted hyperoxia induced downregulation of the expression of genes related to mitotic spindle formation in the WT which was not observed in Sphk1 -/- neonates. Our data clearly suggests a role for SphK1 in neonatal hyperoxic lung injury through elevated inflammation and apoptosis in lung tissue. Further, validation by RT-PCR on 24 differentially expressed genes showed 83% concordance both in terms of fold change and vectorial changes. Our findings are in agreement with previously reported human BPD microarray data and completely support our published in vivo findings. In addition, the data also revealed a significant role for additional unanticipitated signaling pathways involving Wnt and GADD45.
Conclusion: Using SphK1 knockout mice and differential gene expression analysis, we have shown here that S1P/SphK1 signaling plays a key role in promoting hyperoxia induced DNA damage, inflammation, apoptosis and ECM remodeling in neonatal lungs. It also appears to suppress pro-survival cellular responses involved in normal lung development. We therefore propose SphK1 as a therapeutic target for the development drugs to combat BPD.
Keywords: Lipid signaling; Neonatal lung injury; Oxidative stress; Sphingosine 1 phosphate; Sphingosine kinase 1.
Conflict of interest statement
Ethics approval and consent to participate
Not applicable as this is not a study involving human beings. All experiments using animals were approved by the Institutional Animal Care and Use Committee at the University of Illinois at Chicago (protocol # 15-240).
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures
Similar articles
-
The Role of Sphingolipid Signaling in Oxidative Lung Injury and Pathogenesis of Bronchopulmonary Dysplasia.Int J Mol Sci. 2022 Jan 23;23(3):1254. doi: 10.3390/ijms23031254. Int J Mol Sci. 2022. PMID: 35163176 Free PMC article. Review.
-
Sphingosine kinase 1 deficiency confers protection against hyperoxia-induced bronchopulmonary dysplasia in a murine model: role of S1P signaling and Nox proteins.Am J Pathol. 2013 Oct;183(4):1169-1182. doi: 10.1016/j.ajpath.2013.06.018. Epub 2013 Aug 8. Am J Pathol. 2013. PMID: 23933064 Free PMC article.
-
Neonatal therapy with PF543, a sphingosine kinase 1 inhibitor, ameliorates hyperoxia-induced airway remodeling in a murine model of bronchopulmonary dysplasia.Am J Physiol Lung Cell Mol Physiol. 2020 Sep 1;319(3):L497-L512. doi: 10.1152/ajplung.00169.2020. Epub 2020 Jul 22. Am J Physiol Lung Cell Mol Physiol. 2020. PMID: 32697651 Free PMC article.
-
Hyperoxia-induced S1P1 signaling reduced angiogenesis by suppression of TIE-2 leading to experimental bronchopulmonary dysplasia.Cell Biochem Biophys. 2021 Sep;79(3):561-573. doi: 10.1007/s12013-021-01014-8. Epub 2021 Jun 27. Cell Biochem Biophys. 2021. PMID: 34176100 Free PMC article.
-
Epigenetic regulation of pro-inflammatory cytokine secretion by sphingosine 1-phosphate (S1P) in acute lung injury: Role of S1P lyase.Adv Biol Regul. 2017 Jan;63:156-166. doi: 10.1016/j.jbior.2016.09.007. Epub 2016 Sep 29. Adv Biol Regul. 2017. PMID: 27720306 Free PMC article. Review.
Cited by
-
Genetic deletion of Sphk2 confers protection against Pseudomonas aeruginosa mediated differential expression of genes related to virulent infection and inflammation in mouse lung.BMC Genomics. 2019 Dec 16;20(1):984. doi: 10.1186/s12864-019-6367-9. BMC Genomics. 2019. PMID: 31842752 Free PMC article.
-
Investigating the regulatory role of ORMDL3 in airway barrier dysfunction using in vivo and in vitro models.Int J Mol Med. 2019 Aug;44(2):535-548. doi: 10.3892/ijmm.2019.4233. Epub 2019 Jun 6. Int J Mol Med. 2019. PMID: 31173170 Free PMC article.
-
The Role of Sphingolipid Signaling in Oxidative Lung Injury and Pathogenesis of Bronchopulmonary Dysplasia.Int J Mol Sci. 2022 Jan 23;23(3):1254. doi: 10.3390/ijms23031254. Int J Mol Sci. 2022. PMID: 35163176 Free PMC article. Review.
-
Advancements in understanding the role of lysophospholipids and their receptors in lung disorders including bronchopulmonary dysplasia.Biochim Biophys Acta Mol Cell Biol Lipids. 2020 Jul;1865(7):158685. doi: 10.1016/j.bbalip.2020.158685. Epub 2020 Mar 10. Biochim Biophys Acta Mol Cell Biol Lipids. 2020. PMID: 32169655 Free PMC article. Review.
-
Resveratrol alleviates alveolar epithelial cell injury induced by hyperoxia by reducing apoptosis and mitochondrial dysfunction.Exp Biol Med (Maywood). 2021 Mar;246(5):596-606. doi: 10.1177/1535370220975106. Epub 2020 Nov 20. Exp Biol Med (Maywood). 2021. PMID: 33215523 Free PMC article.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
