

Cholesterol metabolism and Cx43, Cx46, and Cx50 gap junction protein expression and localization in normal and diabetic and obese ob/ob and db/db mouse testes
- PMID: 28851737
- PMCID: PMC5866387
- DOI: 10.1152/ajpendo.00215.2017
Cholesterol metabolism and Cx43, Cx46, and Cx50 gap junction protein expression and localization in normal and diabetic and obese ob/ob and db/db mouse testes
Abstract
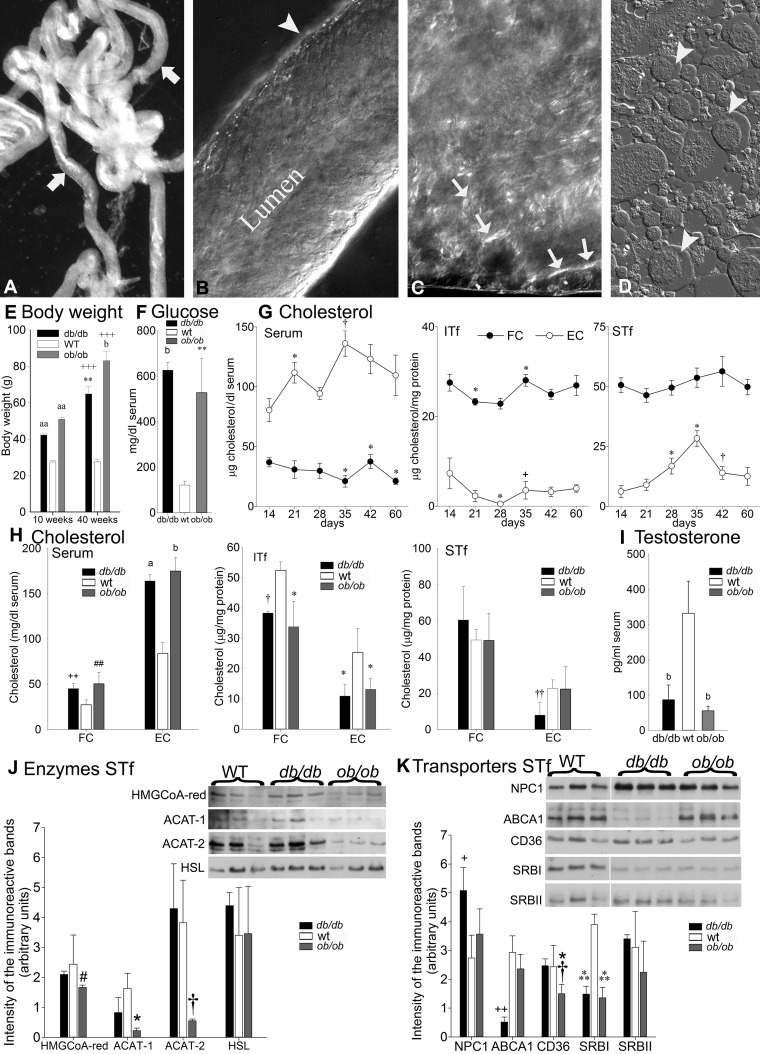
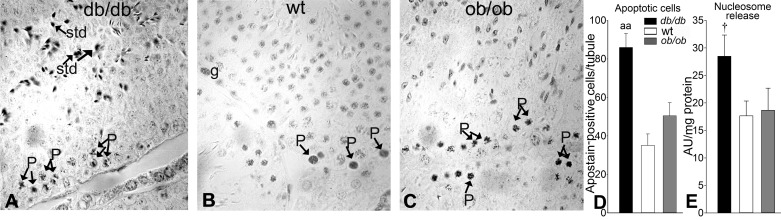
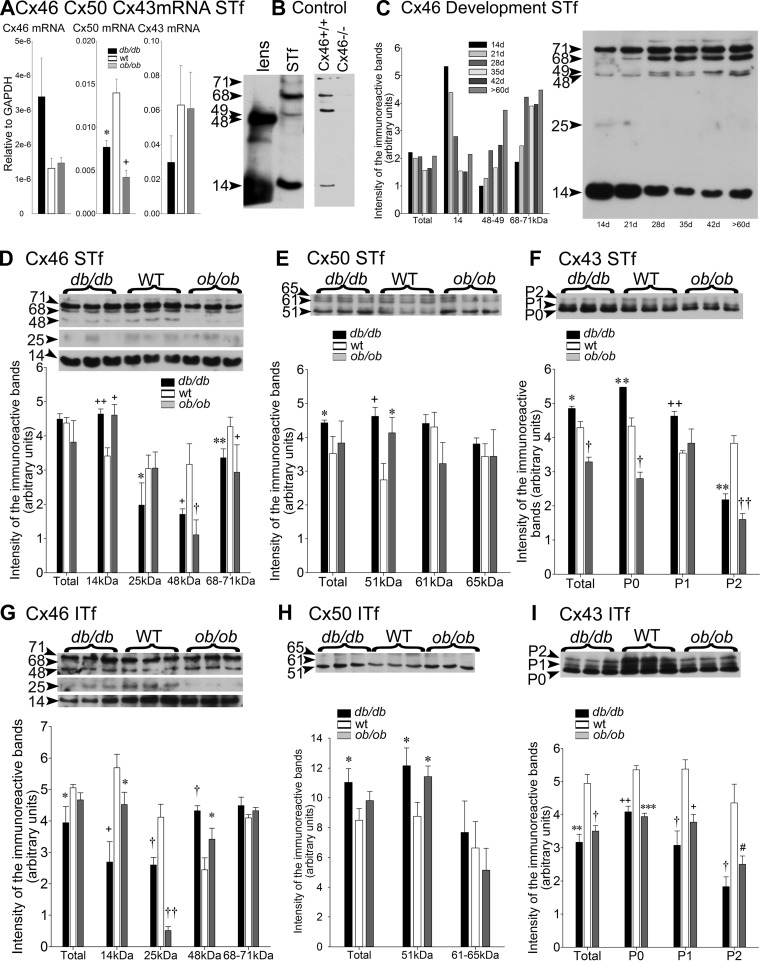
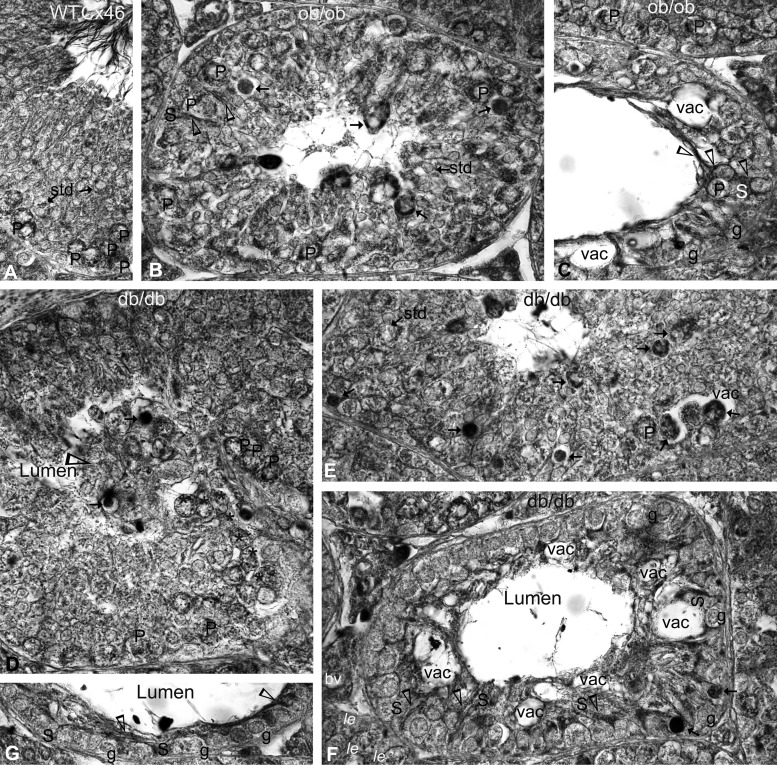
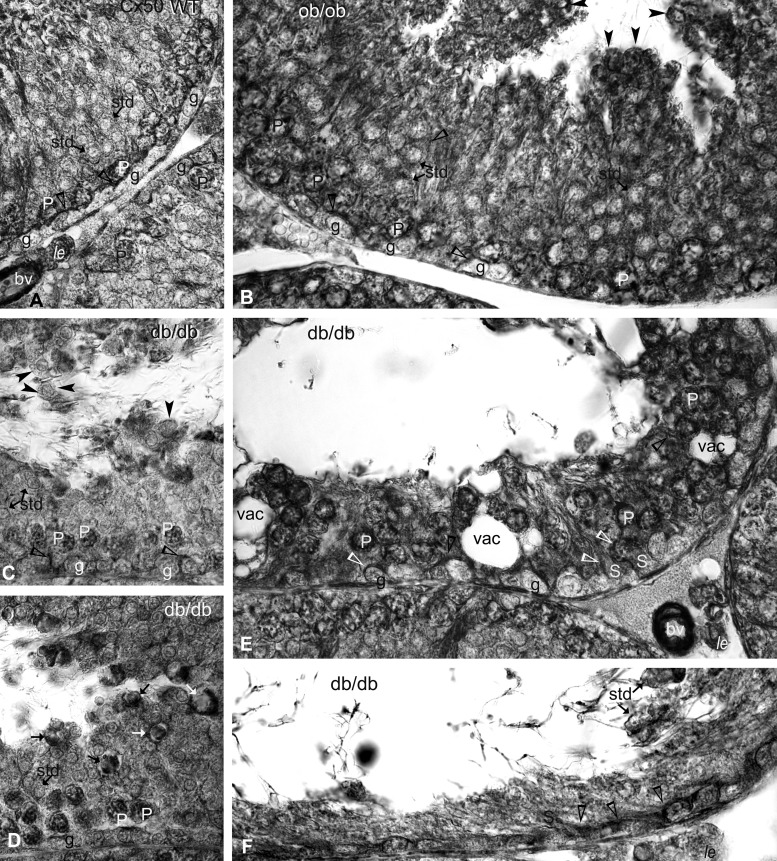
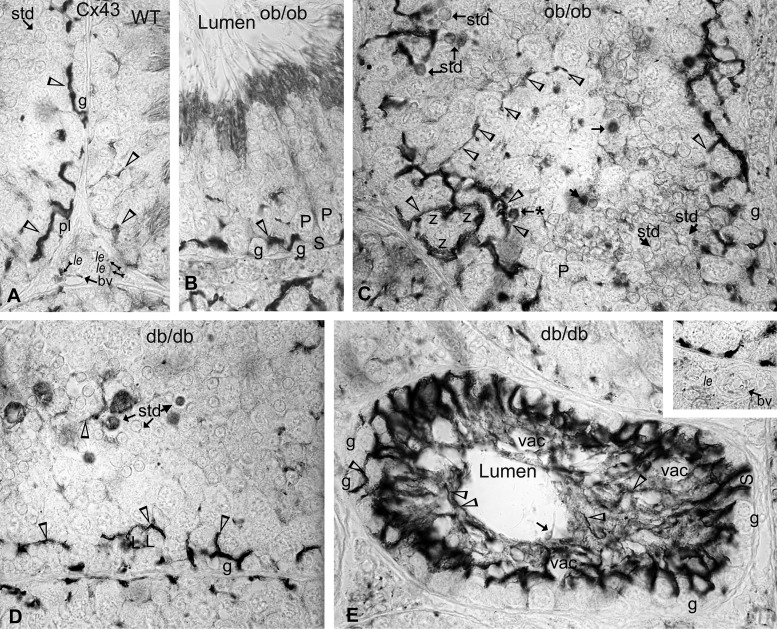
Decreased fertility and birth rates arise from metabolic disorders. This study assesses cholesterol metabolism and Cx46, Cx50, and Cx43 expression in interstitium- and seminiferous tubule-enriched fractions of leptin-deficient ( ob/ob) and leptin receptor-deficient ( db/db) mice, two type 2 diabetes and obesity models associated with infertility. Testosterone levels decreased and glucose and free and esterified cholesterol (FC and EC) levels increased in serum, whereas FC and EC levels decreased in the interstitium, in ob/ob and db/db mice. In tubules, a decrease in EC caused FC-to-EC ratios to increase in db/db mice. In tubules, only acyl coenzyme A:cholesterol acyl transferase type 1 and 2 protein levels significantly decreased in ob/ob, but not db/db, mice compared with wild-type mice, and imbalances in the cholesterol transporters Niemann-Pick C1 (NPC1), ATP-binding cassette A1 (ABCA1), scavenger receptor class B member I (SR-BI), and cluster of differentiation 36 (CD36) were observed in ob/ob and db/db mice. In tubules, 14-kDa Cx46 prevailed during development, 48- to 49- and 68- to 71-kDa Cx46 prevailed during adulthood, and total Cx46 changed little. Compared with wild-type mice, 14-kDa Cx46 increased, whereas 48- to 49- and 68- to 71-kDa Cx46 decreased, in tubules, whereas the opposite occurred in the interstitium, in db/db and ob/ob mice. Total and 51-kDa Cx50 increased in db/db and ob/ob interstitium and tubules. Cx43 levels decreased in ob/ob interstitium and tubules, whereas Cx43 decreased in db/db interstitium but increased in db/db tubules. Apoptosis levels measured by ELISA and numbers of apostain-labeled apoptotic cells significantly increased in db/db, but not ob/ob, tubules. Testicular db/db capillaries were Cx50-positive but weakly Cx43-positive with a thickened lamina, suggesting altered permeability. Our findings indicate that the db mutation-induced impairment of meiosis may arise from imbalances in cholesterol metabolism and upregulated Cx43 expression and phosphorylation in tubules.
Keywords: Sertoli cell; cholesterol; meiosis; spermatogenesis.
Figures
Similar articles
-
Glucose, insulin, insulin receptor subunits α and β in normal and spontaneously diabetic and obese ob/ob and db/db infertile mouse testis and hypophysis.Reprod Biol Endocrinol. 2020 Mar 17;18(1):25. doi: 10.1186/s12958-020-00583-2. Reprod Biol Endocrinol. 2020. PMID: 32183843 Free PMC article.
-
Cx30.2 deletion causes imbalances in testicular Cx43, Cx46, and Cx50 and insulin receptors. Reciprocally, diabetes/obesity alters Cx30.2 in mouse testis.Am J Physiol Regul Integr Comp Physiol. 2020 Jun 1;318(6):R1078-R1090. doi: 10.1152/ajpregu.00044.2020. Epub 2020 Apr 29. Am J Physiol Regul Integr Comp Physiol. 2020. PMID: 32348681 Free PMC article.
-
Complementary expression and phosphorylation of Cx46 and Cx50 during development and following gene deletion in mouse and in normal and orchitic mink testes.Am J Physiol Regul Integr Comp Physiol. 2015 Aug 1;309(3):R255-76. doi: 10.1152/ajpregu.00152.2015. Epub 2015 May 27. Am J Physiol Regul Integr Comp Physiol. 2015. PMID: 26017495 Free PMC article.
-
Mutations of CX46/CX50 and Cataract Development.Front Mol Biosci. 2022 Feb 11;9:842399. doi: 10.3389/fmolb.2022.842399. eCollection 2022. Front Mol Biosci. 2022. PMID: 35223995 Free PMC article. Review.
-
Leptin- and leptin receptor-deficient rodent models: relevance for human type 2 diabetes.Curr Diabetes Rev. 2014 Mar;10(2):131-45. doi: 10.2174/1573399810666140508121012. Curr Diabetes Rev. 2014. PMID: 24809394 Free PMC article. Review.
Cited by
-
High fat diet-induced obesity prolongs critical stages of the spermatogenic cycle in a Ldlr-/-.Leiden mouse model.Sci Rep. 2022 Jan 11;12(1):430. doi: 10.1038/s41598-021-04069-y. Sci Rep. 2022. PMID: 35017550 Free PMC article.
-
Glucose, insulin, insulin receptor subunits α and β in normal and spontaneously diabetic and obese ob/ob and db/db infertile mouse testis and hypophysis.Reprod Biol Endocrinol. 2020 Mar 17;18(1):25. doi: 10.1186/s12958-020-00583-2. Reprod Biol Endocrinol. 2020. PMID: 32183843 Free PMC article.
-
Connexin46 in the nucleus of cancer cells: a possible role as transcription modulator.Cell Commun Signal. 2025 Mar 27;23(1):153. doi: 10.1186/s12964-025-02151-w. Cell Commun Signal. 2025. PMID: 40148950 Free PMC article.
-
What do we know about blood-testis barrier? current understanding of its structure and physiology.Front Cell Dev Biol. 2023 Jun 15;11:1114769. doi: 10.3389/fcell.2023.1114769. eCollection 2023. Front Cell Dev Biol. 2023. PMID: 37397257 Free PMC article. Review.
-
PCSK9 Contributes to the Cholesterol, Glucose, and Insulin2 Homeostasis in Seminiferous Tubules and Maintenance of Immunotolerance in Testis.Front Cell Dev Biol. 2022 May 2;10:889972. doi: 10.3389/fcell.2022.889972. eCollection 2022. Front Cell Dev Biol. 2022. PMID: 35586340 Free PMC article.
References
-
- Akpovi CD, Anago EAA, Segbo JAG, Manindji C, Medehouenou TCM, Loko F, Vitale ML, Pelletier R-M. Blood biochemical parameter levels vary with spermatogenesis in seasonal reproductive model, the mink (Mustela vison). Int J Biosci 6: 222–229, 2015. doi:10.12692/ijb/6.2.222-229. - DOI
-
- Akpovi CD, Pelletier R-M. A revised and improved method for the isolation of seminiferous tubule-enriched fractions that preserves the phosphorylated and glycosylated forms of proteins. In: Human Embryogenesis: Methods and Protocols, edited by Lafond J, Vaillancourt C. Totowa, NJ: Humana Press Springer Protocols, 2009, p. 159–168. doi:10.1007/978-1-60327-009-0_9. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous