

Phosphoproteomic Analysis Reveals the Importance of Kinase Regulation During Orbivirus Infection
- PMID: 28851738
- PMCID: PMC5672004
- DOI: 10.1074/mcp.M117.067355
Phosphoproteomic Analysis Reveals the Importance of Kinase Regulation During Orbivirus Infection
Abstract
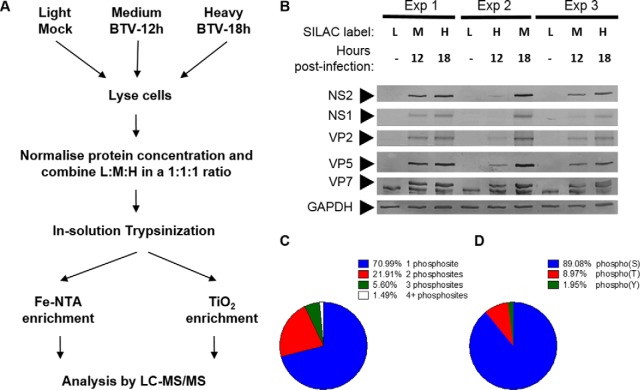
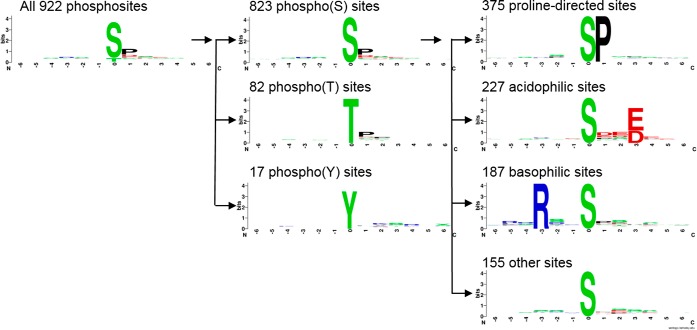
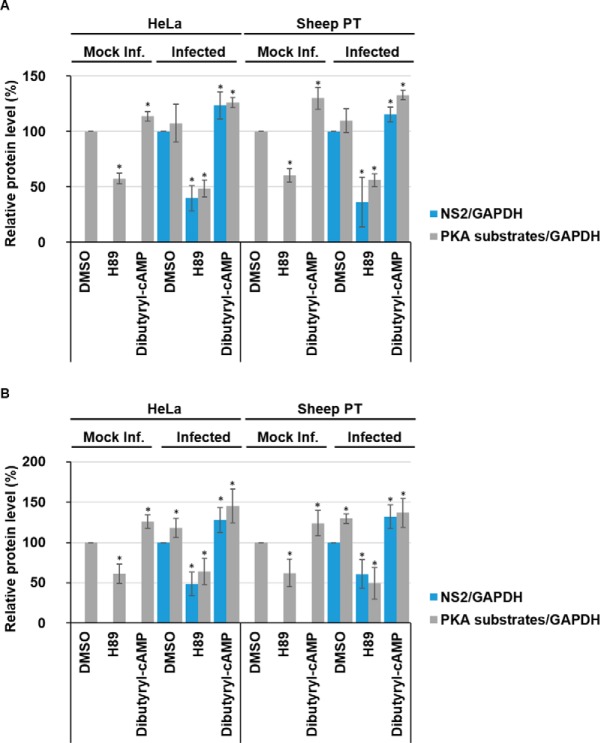
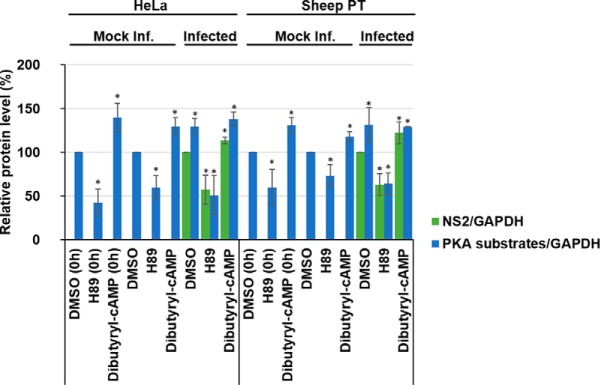
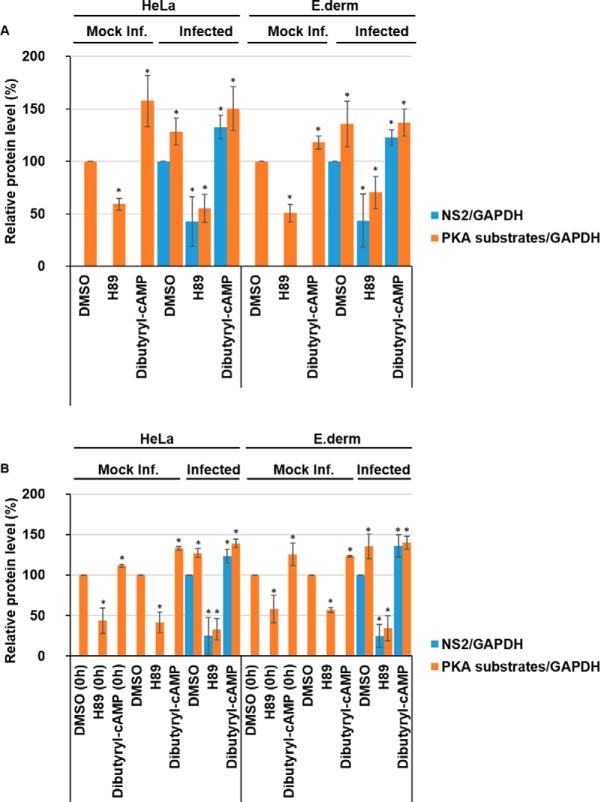
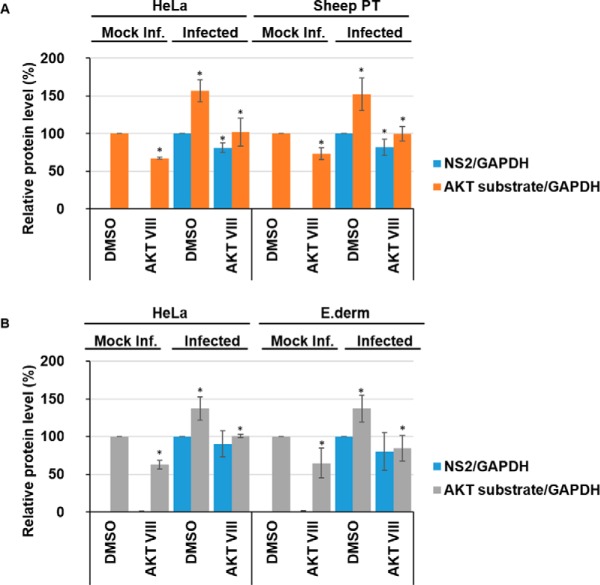
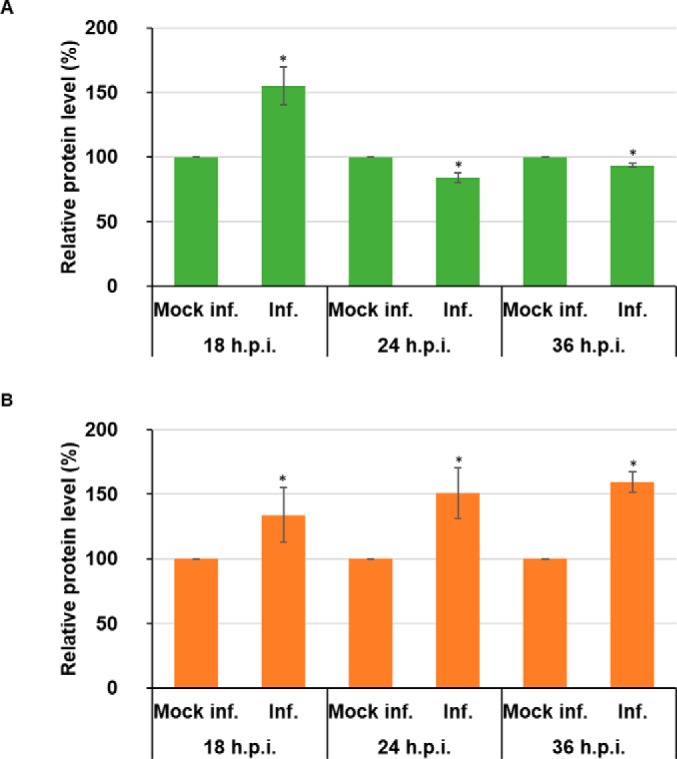
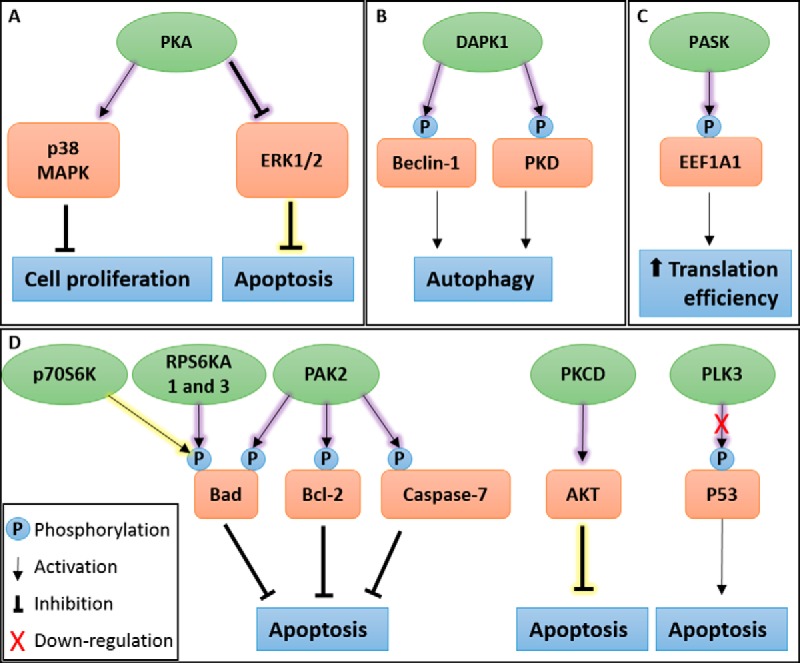
Bluetongue virus (BTV) causes infections in wild and domesticated ruminants with high morbidity and mortality and is responsible for significant economic losses in both developing and developed countries. BTV serves as a model for the study of other members of the Orbivirus genus. Previously, the importance of casein kinase 2 for BTV replication was demonstrated. To identify intracellular signaling pathways and novel host-cell kinases involved during BTV infection, the phosphoproteome of BTV infected cells was analyzed. Over 1000 phosphosites were identified using mass spectrometry, which were then used to determine the corresponding kinases involved during BTV infection. This analysis yielded protein kinase A (PKA) as a novel kinase activated during BTV infection. Subsequently, the importance of PKA for BTV infection was validated using a PKA inhibitor and activator. Our data confirmed that PKA was essential for efficient viral growth. Further, we showed that PKA is also required for infection of equid cells by African horse sickness virus, another member of the Orbivirus genus. Thus, despite their preference in specific host species, orbiviruses may utilize the same host signaling pathways during their replication.
© 2017 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
Similar articles
-
Novel Function of Bluetongue Virus NS3 Protein in Regulation of the MAPK/ERK Signaling Pathway.J Virol. 2019 Jul 30;93(16):e00336-19. doi: 10.1128/JVI.00336-19. Print 2019 Aug 15. J Virol. 2019. PMID: 31167915 Free PMC article.
-
MicroRNA expression profiling of primary sheep testicular cells in response to bluetongue virus infection.Infect Genet Evol. 2017 Apr;49:256-267. doi: 10.1016/j.meegid.2017.01.029. Epub 2017 Jan 27. Infect Genet Evol. 2017. PMID: 28132926
-
Dual modulation of type I interferon response by bluetongue virus.J Virol. 2014 Sep;88(18):10792-802. doi: 10.1128/JVI.01235-14. Epub 2014 Jul 9. J Virol. 2014. PMID: 25008919 Free PMC article.
-
Interferon α/β receptor knockout mice as a model to study bluetongue virus infection.Virus Res. 2014 Mar;182:35-42. doi: 10.1016/j.virusres.2013.09.038. Epub 2013 Oct 4. Virus Res. 2014. PMID: 24100234 Review.
-
Functional mapping of bluetongue virus proteins and their interactions with host proteins during virus replication.Cell Biochem Biophys. 2008;50(3):143-57. doi: 10.1007/s12013-008-9009-4. Epub 2008 Feb 26. Cell Biochem Biophys. 2008. PMID: 18299997 Review.
Cited by
-
Proteomic and Phosphoproteomic Analysis Reveals Differential Immune Response to Hirame Novirhabdovirus (HIRRV) Infection in the Flounder (Paralichthys olivaceus) under Different Temperature.Biology (Basel). 2023 Aug 18;12(8):1145. doi: 10.3390/biology12081145. Biology (Basel). 2023. PMID: 37627029 Free PMC article.
-
CX-4945 Induces Methuosis in Cholangiocarcinoma Cell Lines by a CK2-Independent Mechanism.Cancers (Basel). 2018 Aug 23;10(9):283. doi: 10.3390/cancers10090283. Cancers (Basel). 2018. PMID: 30142881 Free PMC article.
-
Multiple Routes of Bluetongue Virus Egress.Microorganisms. 2020 Jun 27;8(7):965. doi: 10.3390/microorganisms8070965. Microorganisms. 2020. PMID: 32605099 Free PMC article. Review.
-
The alphavirus nonstructural protein 2 NTPase induces a host translational shut-off through phosphorylation of eEF2 via cAMP-PKA-eEF2K signaling.PLoS Pathog. 2023 Feb 27;19(2):e1011179. doi: 10.1371/journal.ppat.1011179. eCollection 2023 Feb. PLoS Pathog. 2023. PMID: 36848386 Free PMC article.
-
Novel Function of Bluetongue Virus NS3 Protein in Regulation of the MAPK/ERK Signaling Pathway.J Virol. 2019 Jul 30;93(16):e00336-19. doi: 10.1128/JVI.00336-19. Print 2019 Aug 15. J Virol. 2019. PMID: 31167915 Free PMC article.
References
-
- Manning G., Whyte D. B., Martinez R., Hunter T., and Sudarsanam S. (2002) The protein kinase complement of the human genome. Science 298, 1912–1934 - PubMed
-
- Modrof J., Muhlberger E., Klenk H. D., and Becker S. (2002) Phosphorylation of VP30 impairs ebola virus transcription. J. Biol. Chem. 277, 33099–33104 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
