

Accessibility explains preferred thiol-disulfide isomerization in a protein domain
- PMID: 28851879
- PMCID: PMC5575259
- DOI: 10.1038/s41598-017-07501-4
Accessibility explains preferred thiol-disulfide isomerization in a protein domain
Abstract
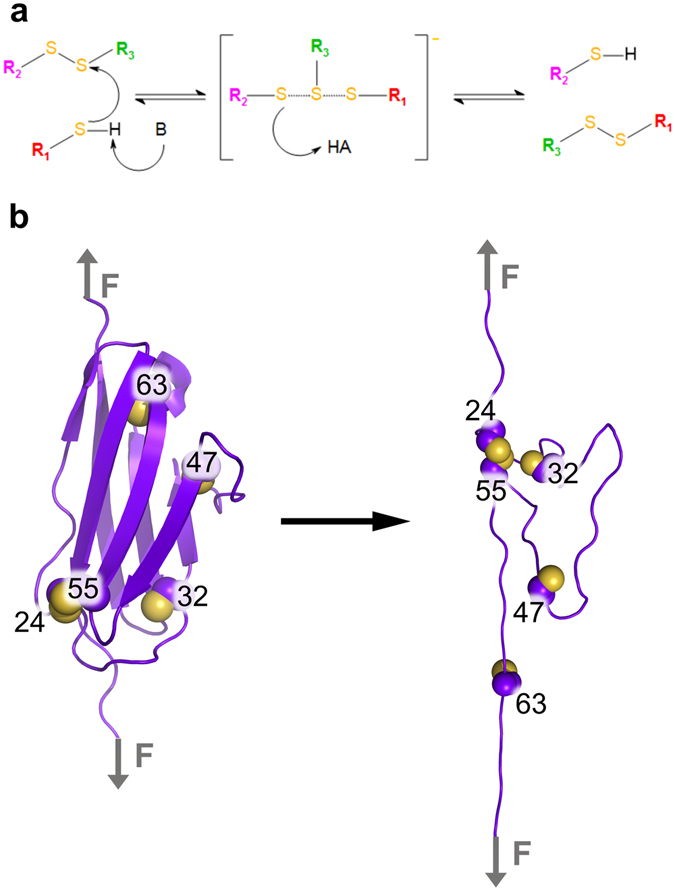
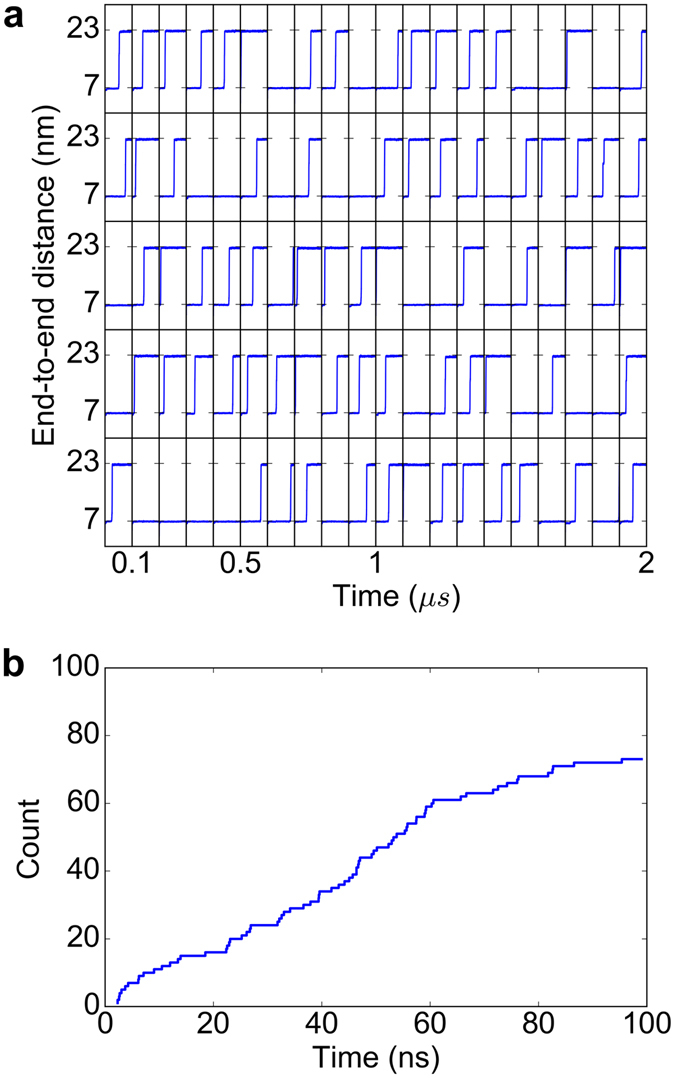
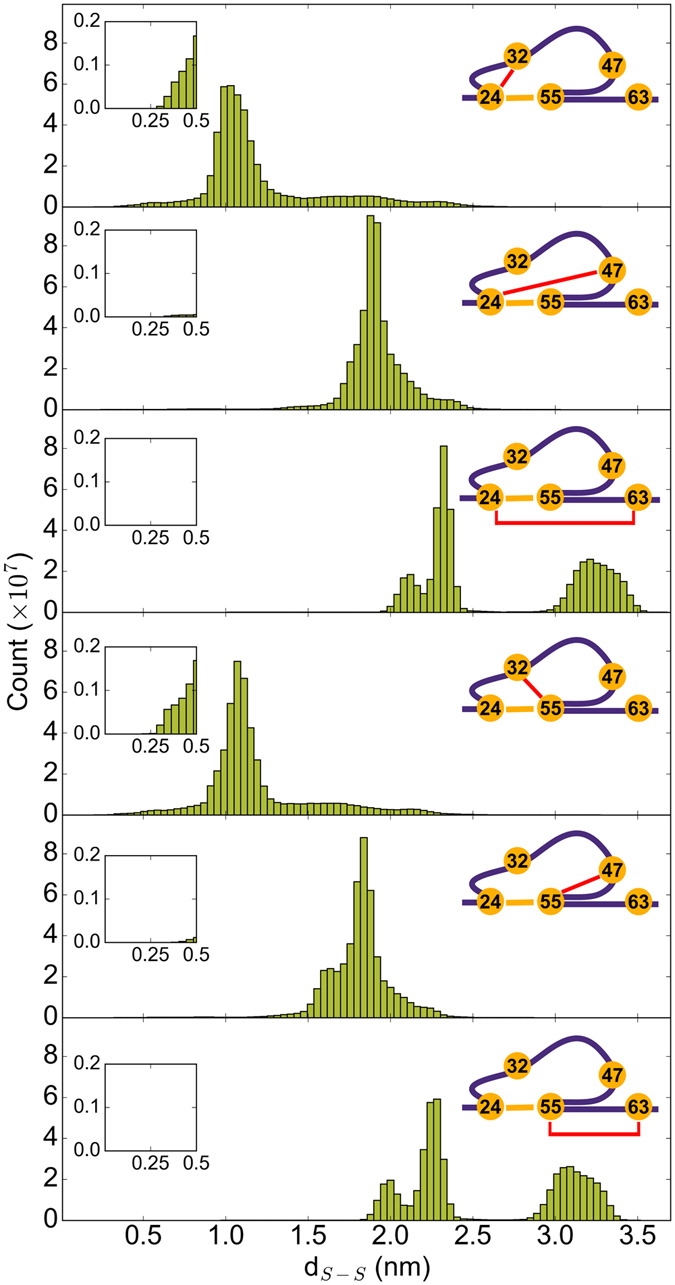
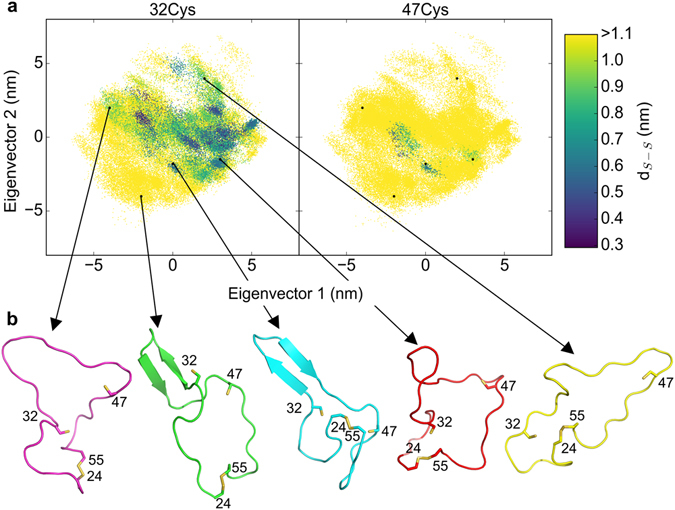
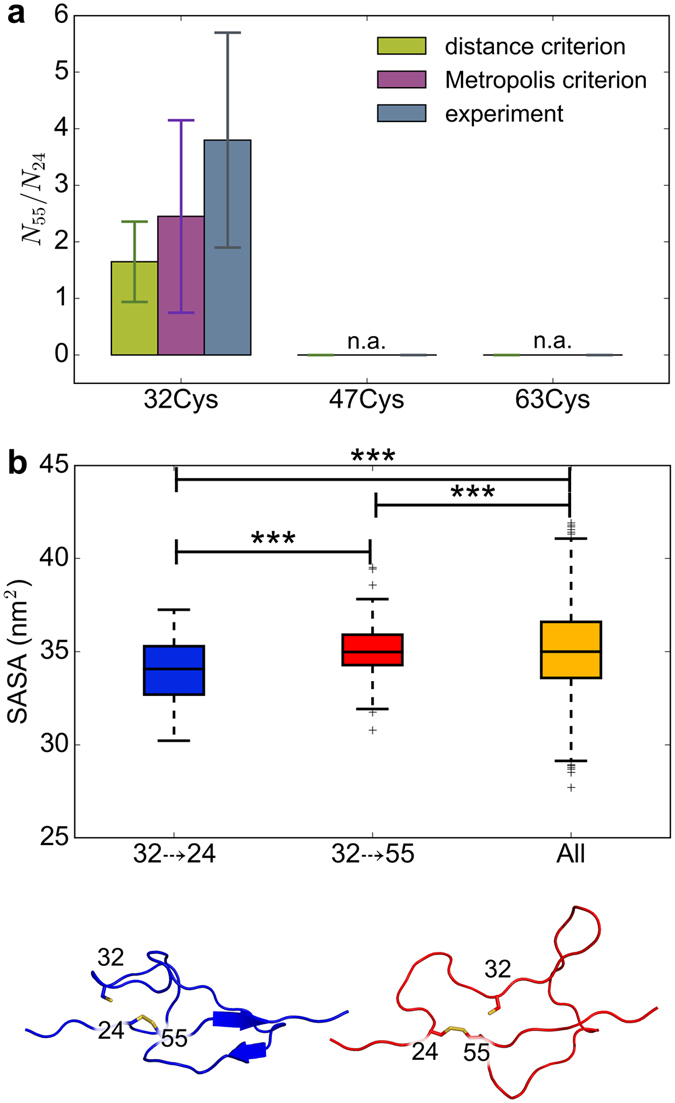
Disulfide bonds are key stabilizing and yet potentially labile cross-links in proteins. While spontaneous disulfide rearrangement through thiol-disulfide exchange is increasingly recognized to play an important physiological role, its molecular determinants are still largely unknown. Here, we used a novel hybrid Monte Carlo and Molecular Dynamics scheme to elucidate the molecular principles of thiol-disulfide exchange in proteins, for a mutated immunoglobulin domain as a model system. Unexpectedly, using simple proximity as the criterion for thiol-disulfide exchange, our method correctly predicts the experimentally observed regiospecificity and selectivity of the cysteine-rich protein. While redox reactivity has been examined primarily on the level of transition states and activation barriers, our results argue for accessibility of the disulfide by the attacking thiol given the highly dynamic and sterically demanding protein as a major bottleneck of thiol-disulfide exchange. This scenario may be similarly at play in other proteins with or without an evolutionarily designed active site.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
