

Retrograde trafficking of β-dystroglycan from the plasma membrane to the nucleus
- PMID: 28852008
- PMCID: PMC5575308
- DOI: 10.1038/s41598-017-09972-x
Retrograde trafficking of β-dystroglycan from the plasma membrane to the nucleus
Erratum in
-
Publisher Correction: Retrograde trafficking of β-dystroglycan from the plasma membrane to the nucleus.Sci Rep. 2018 Dec 7;8(1):17785. doi: 10.1038/s41598-018-36661-0. Sci Rep. 2018. PMID: 30531996 Free PMC article.
Abstract
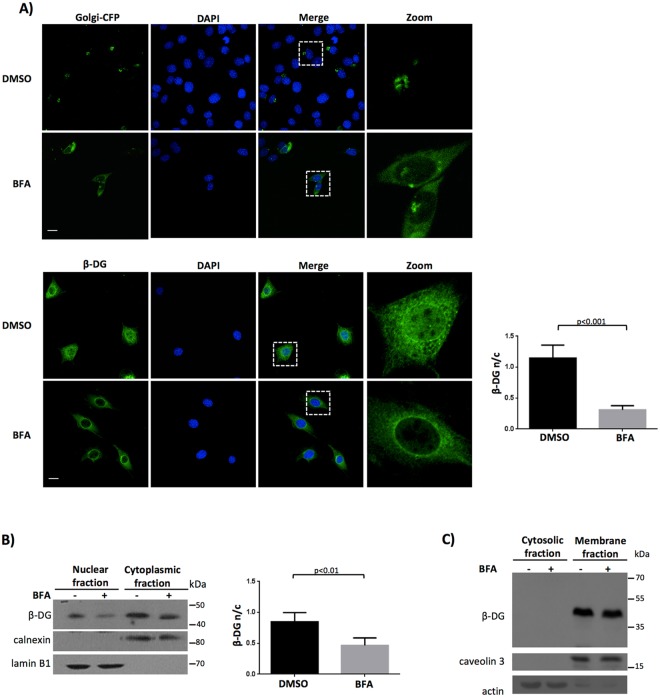
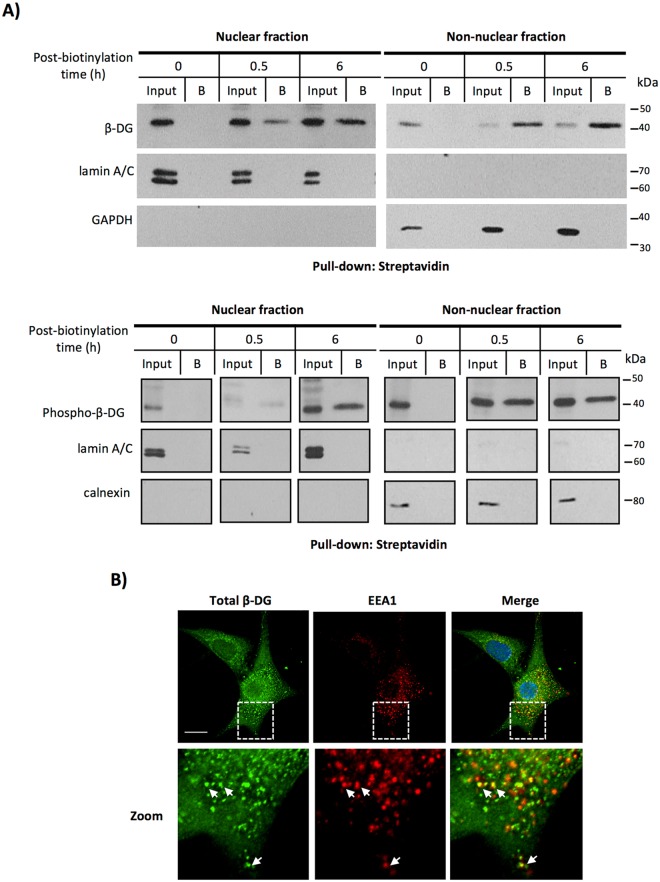
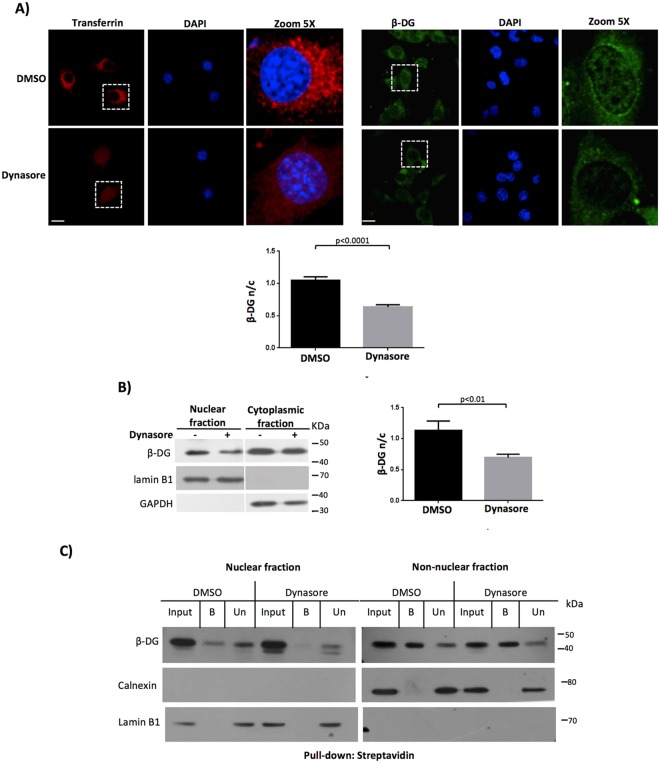
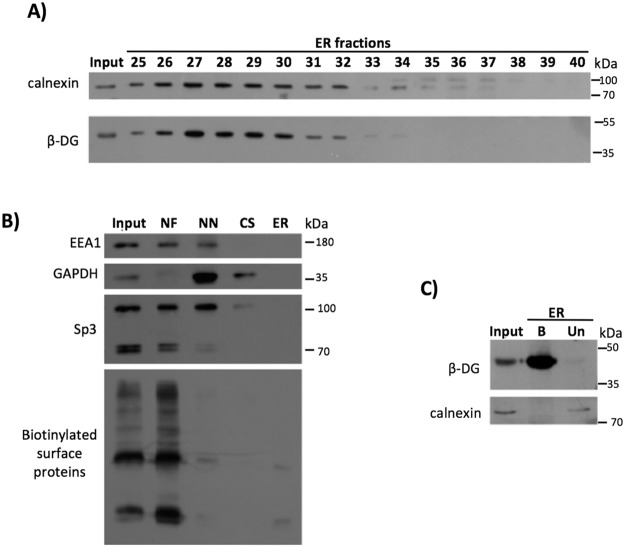
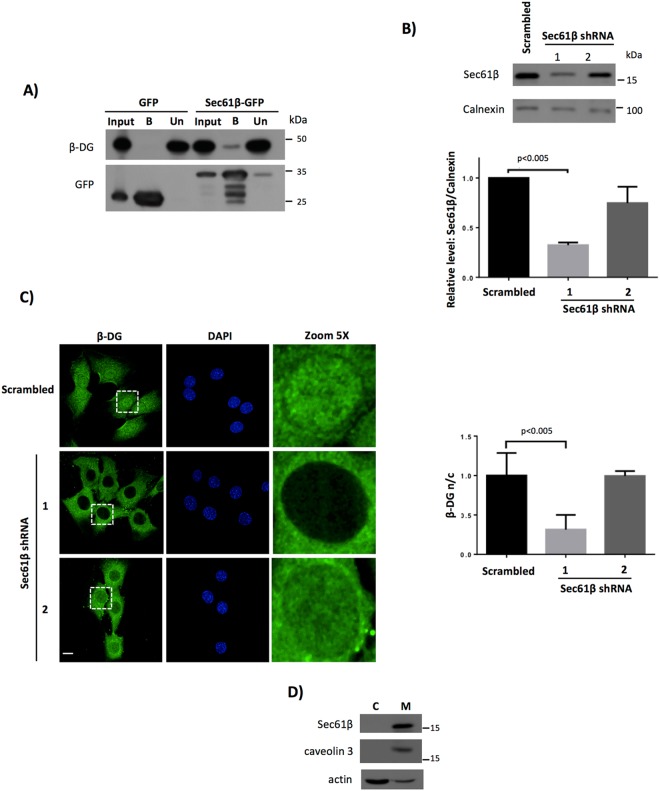
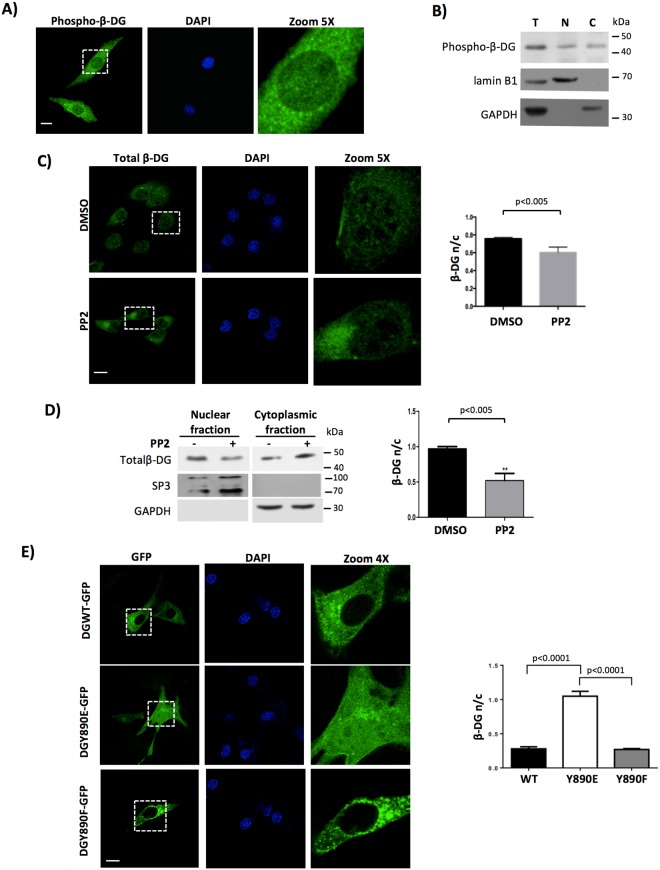
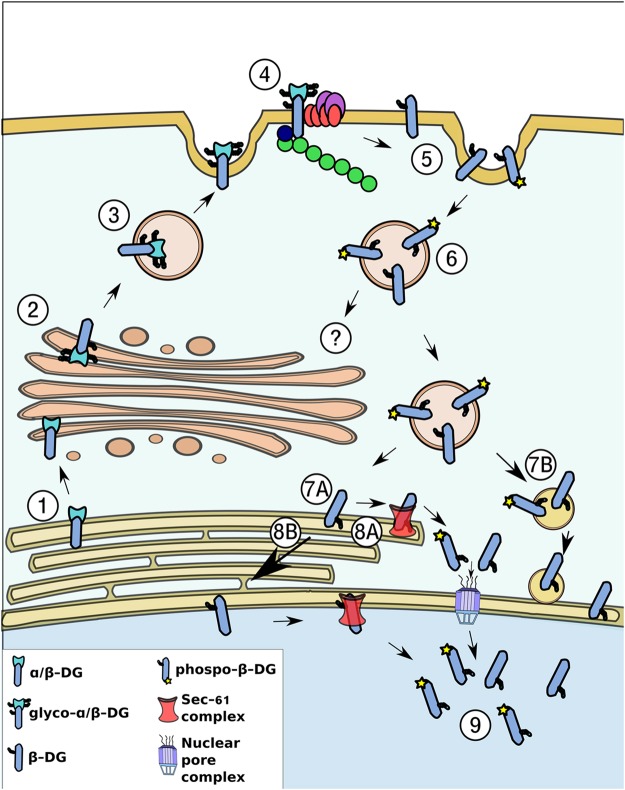
β-Dystroglycan (β-DG) is a transmembrane protein with critical roles in cell adhesion, cytoskeleton remodeling and nuclear architecture. This functional diversity is attributed to the ability of β-DG to target to, and conform specific protein assemblies at the plasma membrane (PM) and nuclear envelope (NE). Although a classical NLS and importin α/β mediated nuclear import pathway has already been described for β-DG, the intracellular trafficking route by which β-DG reaches the nucleus is unknown. In this study, we demonstrated that β-DG undergoes retrograde intracellular trafficking from the PM to the nucleus via the endosome-ER network. Furthermore, we provided evidence indicating that the translocon complex Sec61 mediates the release of β-DG from the ER membrane, making it accessible for importins and nuclear import. Finally, we show that phosphorylation of β-DG at Tyr890 is a key stimulus for β-DG nuclear translocation. Collectively our data describe the retrograde intracellular trafficking route that β-DG follows from PM to the nucleus. This dual role for a cell adhesion receptor permits the cell to functionally connect the PM with the nucleus and represents to our knowledge the first example of a cell adhesion receptor exhibiting retrograde nuclear trafficking and having dual roles in PM and NE.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
Similar articles
-
Characterization of an Importin alpha/beta-recognized nuclear localization signal in beta-dystroglycan.J Cell Biochem. 2010 Jun 1;110(3):706-17. doi: 10.1002/jcb.22581. J Cell Biochem. 2010. PMID: 20512930
-
Nuclear import of β-dystroglycan is facilitated by ezrin-mediated cytoskeleton reorganization.PLoS One. 2014 Mar 5;9(3):e90629. doi: 10.1371/journal.pone.0090629. eCollection 2014. PLoS One. 2014. PMID: 24599031 Free PMC article.
-
Control of nuclear β-dystroglycan content is crucial for the maintenance of nuclear envelope integrity and function.Biochim Biophys Acta Mol Cell Res. 2018 Feb;1865(2):406-420. doi: 10.1016/j.bbamcr.2017.11.013. Epub 2017 Nov 21. Biochim Biophys Acta Mol Cell Res. 2018. PMID: 29175376
-
From adhesion complex to signaling hub: the dual role of dystroglycan.Front Mol Biosci. 2023 Dec 14;10:1325284. doi: 10.3389/fmolb.2023.1325284. eCollection 2023. Front Mol Biosci. 2023. PMID: 38155958 Free PMC article. Review.
-
Nuclear-cytoplasmatic shuttling of proteins in control of cellular oxygen sensing.J Mol Med (Berl). 2015 Jun;93(6):599-608. doi: 10.1007/s00109-015-1276-0. Epub 2015 Mar 27. J Mol Med (Berl). 2015. PMID: 25809665 Review.
Cited by
-
The intracellular domain of β-dystroglycan mediates the nucleolar stress response by suppressing UBF transcriptional activity.Cell Death Dis. 2019 Feb 27;10(3):196. doi: 10.1038/s41419-019-1454-z. Cell Death Dis. 2019. PMID: 30814495 Free PMC article.
-
The Molecular Basis and Biologic Significance of the β-Dystroglycan-Emerin Interaction.Int J Mol Sci. 2020 Aug 19;21(17):5944. doi: 10.3390/ijms21175944. Int J Mol Sci. 2020. PMID: 32824881 Free PMC article.
-
The role of laminins in cancer pathobiology: a comprehensive review.J Transl Med. 2025 Jan 17;23(1):83. doi: 10.1186/s12967-025-06079-0. J Transl Med. 2025. PMID: 39825429 Free PMC article. Review.
-
Profiling of the muscle-specific dystroglycan interactome reveals the role of Hippo signaling in muscular dystrophy and age-dependent muscle atrophy.BMC Med. 2020 Jan 21;18(1):8. doi: 10.1186/s12916-019-1478-3. BMC Med. 2020. PMID: 31959160 Free PMC article.
-
Loss of Dystroglycan Drives Cellular Senescence via Defective Mitosis-Mediated Genomic Instability.Int J Mol Sci. 2020 Jul 14;21(14):4961. doi: 10.3390/ijms21144961. Int J Mol Sci. 2020. PMID: 32674290 Free PMC article.
References
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
