

Pheromones modulate reward responsiveness and non-associative learning in honey bees
- PMID: 28852036
- PMCID: PMC5574997
- DOI: 10.1038/s41598-017-10113-7
Pheromones modulate reward responsiveness and non-associative learning in honey bees
Abstract
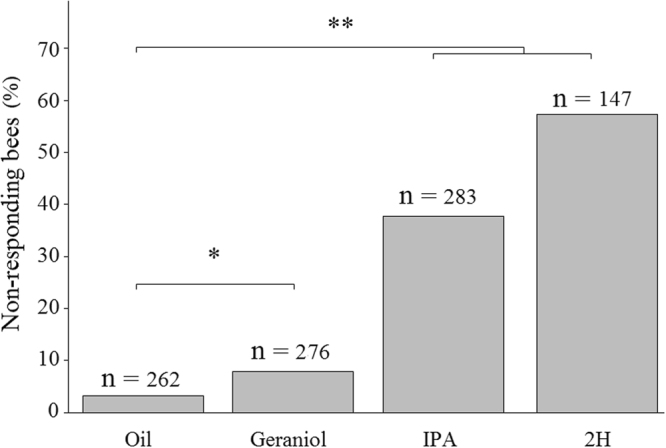
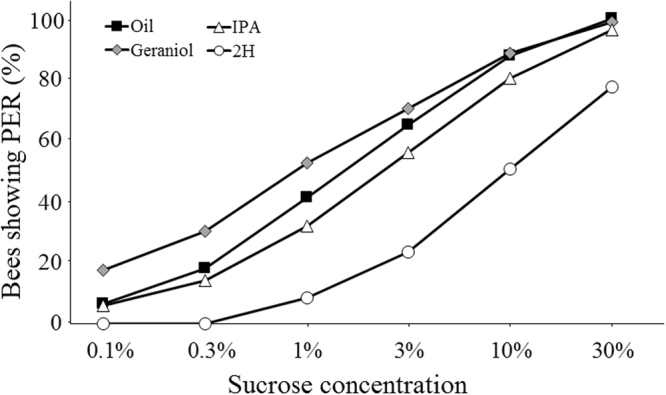
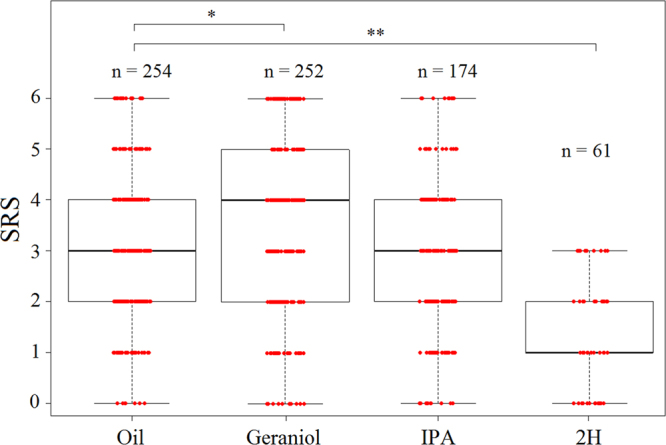
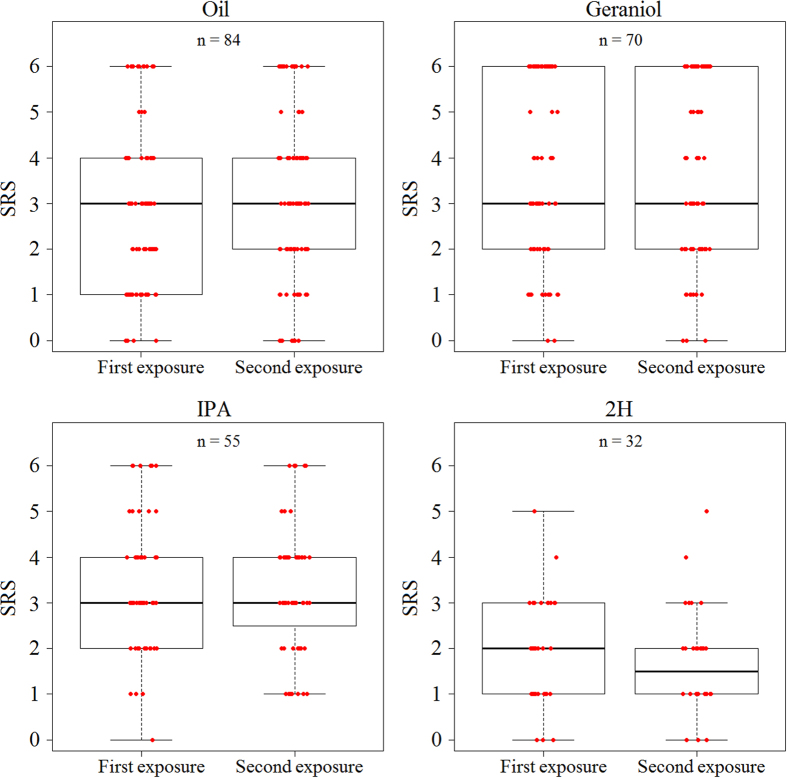
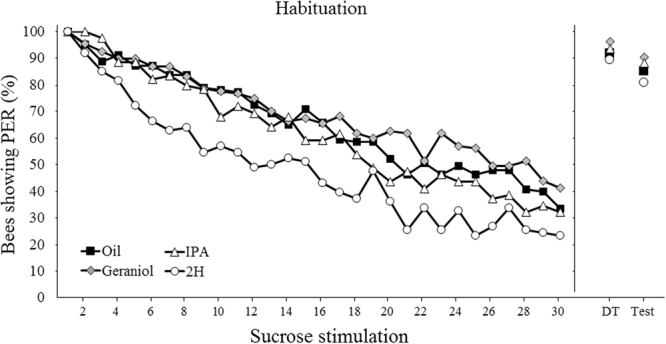
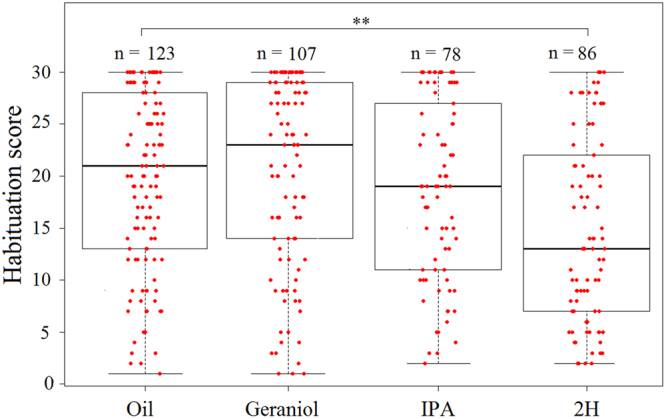
Pheromones are chemical messengers that trigger stereotyped behaviors and/or physiological processes in individuals of the same species. Recent reports suggest that pheromones can modulate behaviors not directly related to the pheromonal message itself and contribute, in this way, to behavioral plasticity. We tested this hypothesis by studying the effect of pheromones on sucrose responsiveness and habituation in honey bees. We exposed workers to three pheromone components: geraniol, which in nature is used in an appetitive context, and isopentyl acetate (IPA) and 2-heptanone (2H), which signal aversive situations. Pheromones associated with an aversive context induced a significant decrease of sucrose responsiveness as 40% and 60% of bees exposed to IPA and 2H, respectively, did not respond to any sucrose concentration. In bees that responded to sucrose, geraniol enhanced sucrose responsiveness while 2H, but not IPA, had the opposite effect. Geraniol and IPA had no effect on habituation while 2H induced faster habituation than controls. Overall, our results demonstrate that pheromones modulate reward responsiveness and to a lower degree habituation. Through their effect on sucrose responsiveness they could also affect appetitive associative learning. Thus, besides conveying stereotyped messages, pheromones may contribute to individual and colony-level plasticity by modulating motivational state and learning performances.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
Similar articles
-
Pheromones modulate responsiveness to a noxious stimulus in honey bees.J Exp Biol. 2018 Mar 13;221(Pt 5):jeb172270. doi: 10.1242/jeb.172270. J Exp Biol. 2018. PMID: 29378816
-
Honey Bees Modulate Their Olfactory Learning in the Presence of Hornet Predators and Alarm Component.PLoS One. 2016 Feb 26;11(2):e0150399. doi: 10.1371/journal.pone.0150399. eCollection 2016. PLoS One. 2016. PMID: 26919132 Free PMC article.
-
Pheromone components affect motivation and induce persistent modulation of associative learning and memory in honey bees.Commun Biol. 2020 Aug 17;3(1):447. doi: 10.1038/s42003-020-01183-x. Commun Biol. 2020. PMID: 32807870 Free PMC article.
-
Bee pheromones: signal or agent of manipulation?Curr Biol. 2009 Jul 28;19(14):R547-8. doi: 10.1016/j.cub.2009.05.032. Curr Biol. 2009. PMID: 19640487 Review.
-
Defensive behavior of honey bees: organization, genetics, and comparisons with other bees.Annu Rev Entomol. 2004;49:271-98. doi: 10.1146/annurev.ento.49.061802.123155. Annu Rev Entomol. 2004. PMID: 14651465 Review.
Cited by
-
Differential Brain Expression Patterns of microRNAs Related to Olfactory Performance in Honey Bees (Apis mellifera).Genes (Basel). 2023 Apr 28;14(5):1000. doi: 10.3390/genes14051000. Genes (Basel). 2023. PMID: 37239360 Free PMC article.
-
Revealing antagonistic interactions in the adverse effects of polystyrene and poly(methyl methacrylate) microplastics in bumblebees.Proc Biol Sci. 2025 Mar;292(2043):20250047. doi: 10.1098/rspb.2025.0047. Epub 2025 Mar 26. Proc Biol Sci. 2025. PMID: 40132636
-
Trail Pheromone Does Not Modulate Subjective Reward Evaluation in Lasius niger Ants.Front Psychol. 2020 Sep 23;11:555576. doi: 10.3389/fpsyg.2020.555576. eCollection 2020. Front Psychol. 2020. PMID: 33071878 Free PMC article.
-
Neuronal Response Latencies Encode First Odor Identity Information across Subjects.J Neurosci. 2018 Oct 24;38(43):9240-9251. doi: 10.1523/JNEUROSCI.0453-18.2018. Epub 2018 Sep 10. J Neurosci. 2018. PMID: 30201774 Free PMC article.
-
Adipokinetic hormone (AKH), energy budget and their effect on feeding and gustatory processes of foraging honey bees.Sci Rep. 2021 Sep 15;11(1):18311. doi: 10.1038/s41598-021-97851-x. Sci Rep. 2021. PMID: 34526585 Free PMC article.
References
-
- Wyatt, T. D. Pheromones and animal behavior: chemical signals and signatures 2nd edition. (Cambridge University Press, 2014).
-
- Seeley, T. D. Honeybee ecology: a study of adaptation in social life (Princeton University Press, 2014).
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
