

Regulation of heat shock transcription factors and their roles in physiology and disease
- PMID: 28852220
- PMCID: PMC5794010
- DOI: 10.1038/nrm.2017.73
Regulation of heat shock transcription factors and their roles in physiology and disease
Abstract
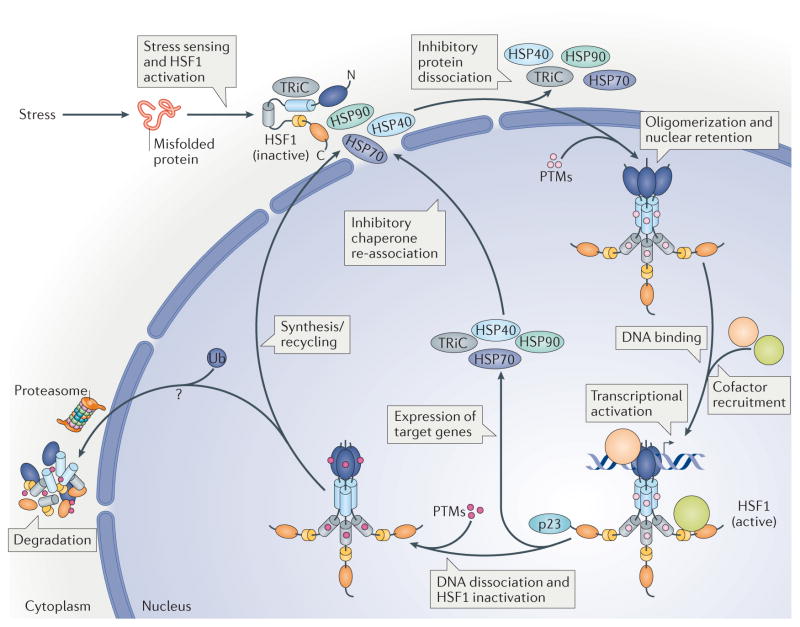
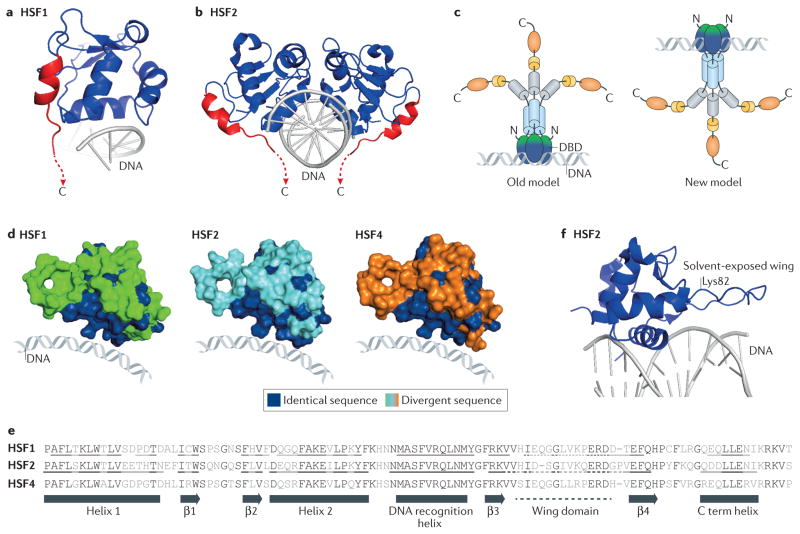
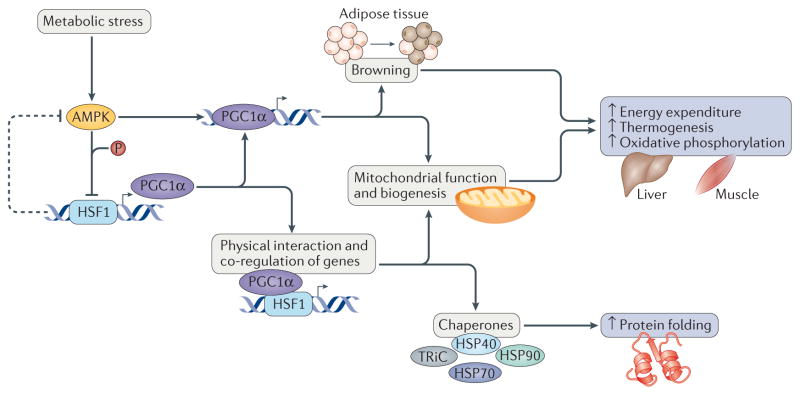
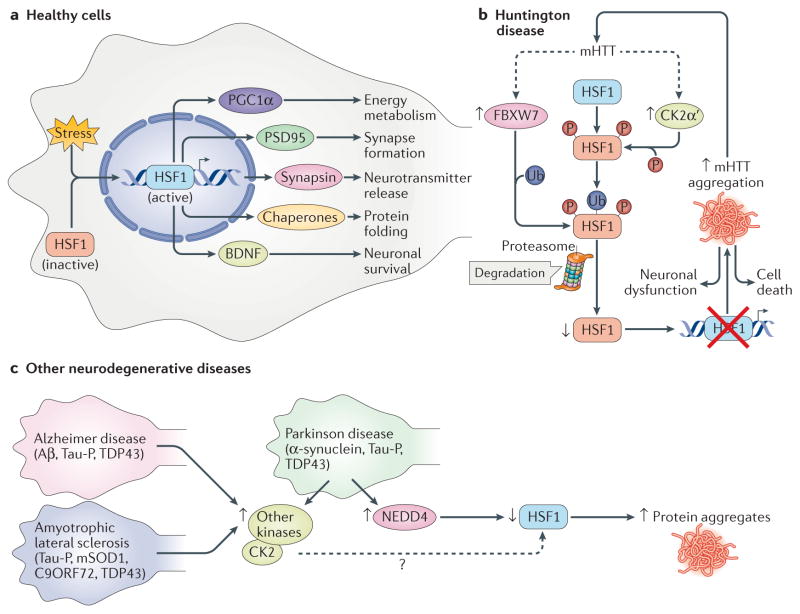
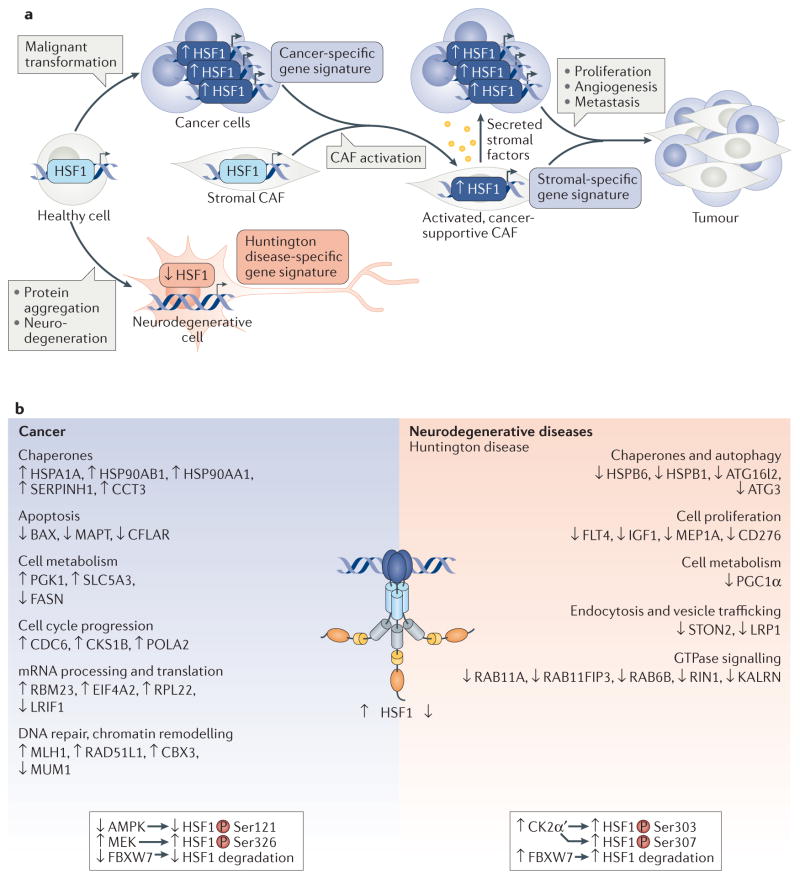
The heat shock transcription factors (HSFs) were discovered over 30 years ago as direct transcriptional activators of genes regulated by thermal stress, encoding heat shock proteins. The accepted paradigm posited that HSFs exclusively activate the expression of protein chaperones in response to conditions that cause protein misfolding by recognizing a simple promoter binding site referred to as a heat shock element. However, we now realize that the mammalian family of HSFs comprises proteins that independently or in concert drive combinatorial gene regulation events that activate or repress transcription in different contexts. Advances in our understanding of HSF structure, post-translational modifications and the breadth of HSF-regulated target genes have revealed exciting new mechanisms that modulate HSFs and shed new light on their roles in physiology and pathology. For example, the ability of HSF1 to protect cells from proteotoxicity and cell death is impaired in neurodegenerative diseases but can be exploited by cancer cells to support their growth, survival and metastasis. These new insights into HSF structure, function and regulation should facilitate the development tof new disease therapeutics to manipulate this transcription factor family.
Conflict of interest statement
The authors declare competing interests: see Web version for details.
Figures
References
-
- Anckar J, Sistonen L. Regulation of HSF1 function in the heat stress response: implications in aging and disease. Annu Rev Biochem. 2011;80:1089–1115. - PubMed
-
- Gomez-Pastor R, et al. Abnormal degradation of the neuronal stress-protective transcription factor HSF1 in Huntington’s disease. Nat Commun. 2017;8:14405. This publication reported a mechanism for the dampened expression of chaperones in polyQ expansion disease through the targeted degradation of HSF1. - PMC - PubMed
-
- Mendillo ML, et al. HSF1 drives a transcriptional program distinct from heat shock to support highly malignant human cancers. Cell. 2012;150:549–562. This work identifies the HSF1 cancer gene signature, a set of genes that are largely distinct from those activated by heat shock stress. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
