

Omics data reveal the unusual asexual-fruiting nature and secondary metabolic potentials of the medicinal fungus Cordyceps cicadae
- PMID: 28854898
- PMCID: PMC5577849
- DOI: 10.1186/s12864-017-4060-4
Omics data reveal the unusual asexual-fruiting nature and secondary metabolic potentials of the medicinal fungus Cordyceps cicadae
Abstract
Background: Ascomycete Cordyceps species have been using as valued traditional Chinese medicines. Particularly, the fruiting bodies of Cordyceps cicadae (syn. Isaria cicadae) have long been utilized for the treatment of chronic kidney disease. However, the genetics and bioactive chemicals in this fungus have been largely unexplored.
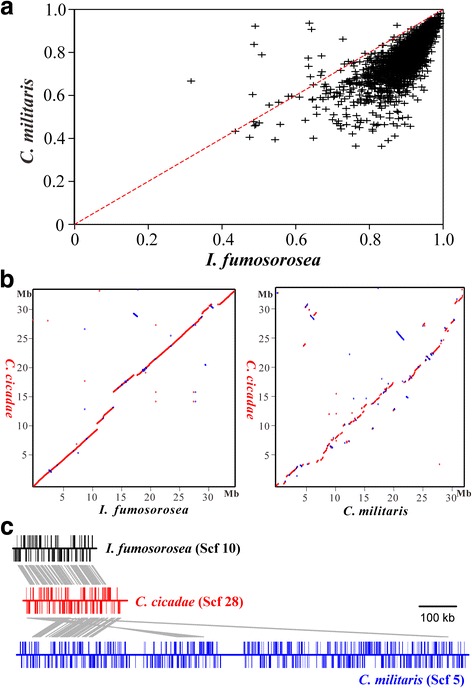
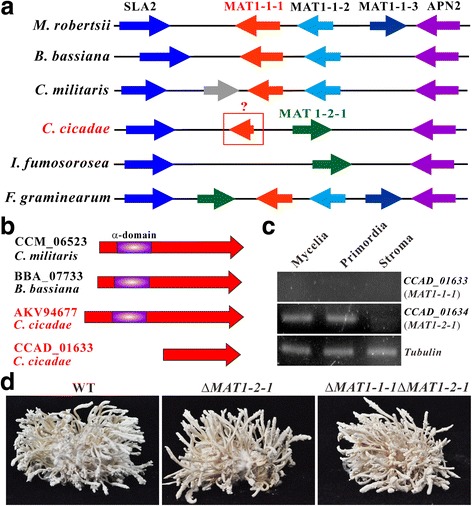
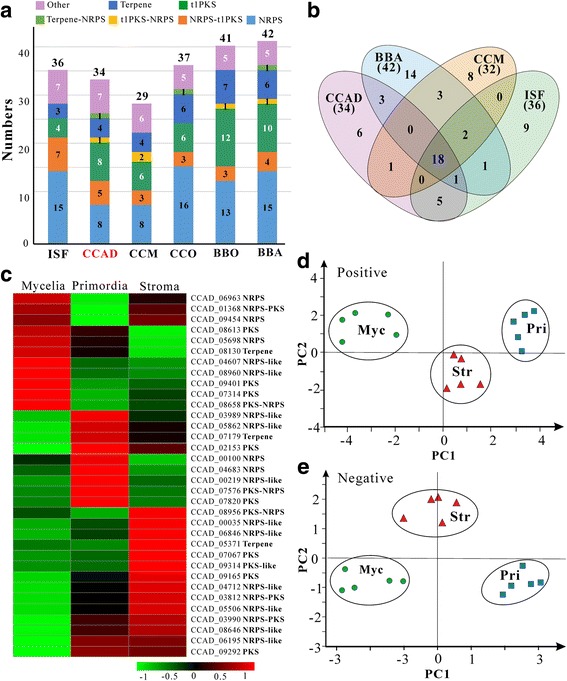
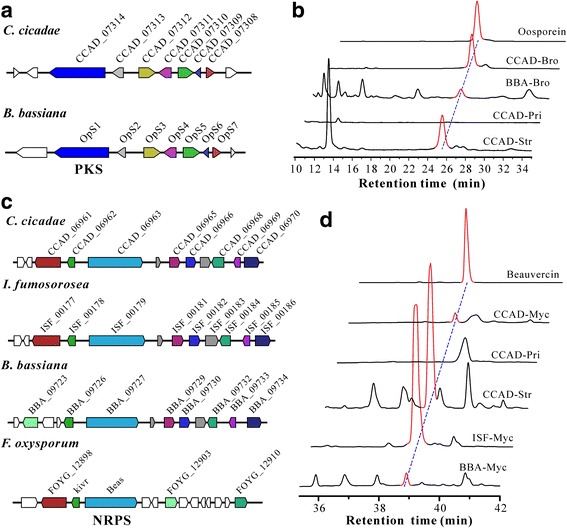
Results: In this study, we performed comprehensive omics analyses of C. cicadae, and found that, in contrast to other Cordyceps fungi, C. cicadae produces asexual fruiting bodies with the production of conidial spores instead of the meiotic ascospores. Genome sequencing and comparative genomic analysis indicate that the protein families encoded by C. cicadae are typical of entomopathogenic fungi, including the expansion of proteases and chitinases for targeting insect hosts. Interestingly, we found that the MAT1-2 mating-type locus of the sequenced strain contains an abnormally truncated MAT1-1-1 gene. Gene deletions revealed that asexual fruiting of C. cicadae is independent of the MAT locus control. RNA-seq transcriptome data also indicate that, compared to growth in a liquid culture, the putative genes involved in mating and meiosis processes were not up-regulated during fungal fruiting, further supporting asexual reproduction in this fungus. The genome of C. cicadae encodes an array of conservative and divergent gene clusters for secondary metabolisms. Based on our analysis, the production of known carcinogenic metabolites by this fungus could be potentially precluded. However, the confirmed production of oosporein raises health concerns about the frequent consumption of fungal fruiting bodies.
Conclusions: The results of this study expand our knowledge of fungal genetics that asexual fruiting can occur independent of the MAT locus control. The obtained genomic and metabolomic data will benefit future investigations of this fungus for medicinal uses.
Keywords: Asexual fruiting; Bioactive metabolites; Cordyceps cicadae; Genomics; Mating type; Secondary metabolism.
Conflict of interest statement
Ethics approval and consent to participate
Field permission is not required for the obtaining of fungal samples in this study.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures

Similar articles
-
Bioactive Metabolites and Potential Mycotoxins Produced by Cordyceps Fungi: A Review of Safety.Toxins (Basel). 2020 Jun 19;12(6):410. doi: 10.3390/toxins12060410. Toxins (Basel). 2020. PMID: 32575649 Free PMC article. Review.
-
Functional convergence and divergence of mating-type genes fulfilling in Cordyceps militaris.Fungal Genet Biol. 2016 Mar;88:35-43. doi: 10.1016/j.fgb.2016.01.013. Epub 2016 Jan 23. Fungal Genet Biol. 2016. PMID: 26812121
-
Genome sequence of the insect pathogenic fungus Cordyceps militaris, a valued traditional Chinese medicine.Genome Biol. 2011 Nov 23;12(11):R116. doi: 10.1186/gb-2011-12-11-r116. Genome Biol. 2011. PMID: 22112802 Free PMC article.
-
Heterothallism in Cordyceps takaomontana.FEMS Microbiol Lett. 2005 Sep 1;250(1):145-50. doi: 10.1016/j.femsle.2005.07.004. FEMS Microbiol Lett. 2005. PMID: 16055279
-
Genetics of Cordyceps and related fungi.Appl Microbiol Biotechnol. 2013 Apr;97(7):2797-804. doi: 10.1007/s00253-013-4771-7. Epub 2013 Feb 23. Appl Microbiol Biotechnol. 2013. PMID: 23435902 Review.
Cited by
-
Secondary metabolites (SMs) of Isaria cicadae and Isaria tenuipes.RSC Adv. 2018 Dec 21;9(1):172-184. doi: 10.1039/c8ra09039d. eCollection 2018 Dec 19. RSC Adv. 2018. PMID: 35521576 Free PMC article. Review.
-
Multi-omics insights into growth and fruiting body development in the entomopathogenic fungus Cordycepsblackwelliae.IMA Fungus. 2025 May 7;16:e147558. doi: 10.3897/imafungus.16.147558. eCollection 2025. IMA Fungus. 2025. PMID: 40376252 Free PMC article.
-
Production of Diverse Beauveriolide Analogs in Closely Related Fungi: a Rare Case of Fungal Chemodiversity.mSphere. 2020 Sep 2;5(5):e00667-20. doi: 10.1128/mSphere.00667-20. mSphere. 2020. PMID: 32878933 Free PMC article.
-
Identification of cordycepin biosynthesis-related genes through de novo transcriptome assembly and analysis in Cordyceps cicadae.R Soc Open Sci. 2018 Dec 19;5(12):181247. doi: 10.1098/rsos.181247. eCollection 2018 Dec. R Soc Open Sci. 2018. PMID: 30662735 Free PMC article.
-
Bioactive Metabolites and Potential Mycotoxins Produced by Cordyceps Fungi: A Review of Safety.Toxins (Basel). 2020 Jun 19;12(6):410. doi: 10.3390/toxins12060410. Toxins (Basel). 2020. PMID: 32575649 Free PMC article. Review.
References
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
