

Immunogenicity of the Plasmodium vivax merozoite surface protein 1 paralog in the induction of naturally acquired antibody and memory B cell responses
- PMID: 28854974
- PMCID: PMC5577667
- DOI: 10.1186/s12936-017-2000-z
Immunogenicity of the Plasmodium vivax merozoite surface protein 1 paralog in the induction of naturally acquired antibody and memory B cell responses
Abstract
Background: The Plasmodium vivax merozoite surface protein 1 paralog (PvMSP1P-19) is a glycosylphosphatidylinositol (GPI)-anchored blood-stage protein that is expressed on the merozoite surface. It is proposed as a blood-stage vaccine candidate against P. vivax because of its ability to induce immune responses upon natural P. vivax exposure and in immunized animals. This study aimed to demonstrate the presence of inhibitory antibodies and memory B cell responses to the PvMSP1P-19 antigen during acute P. vivax infection and after recovery from infection.
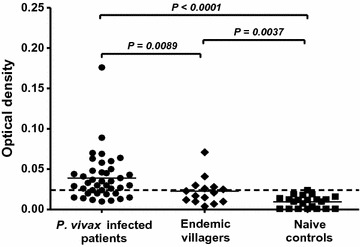
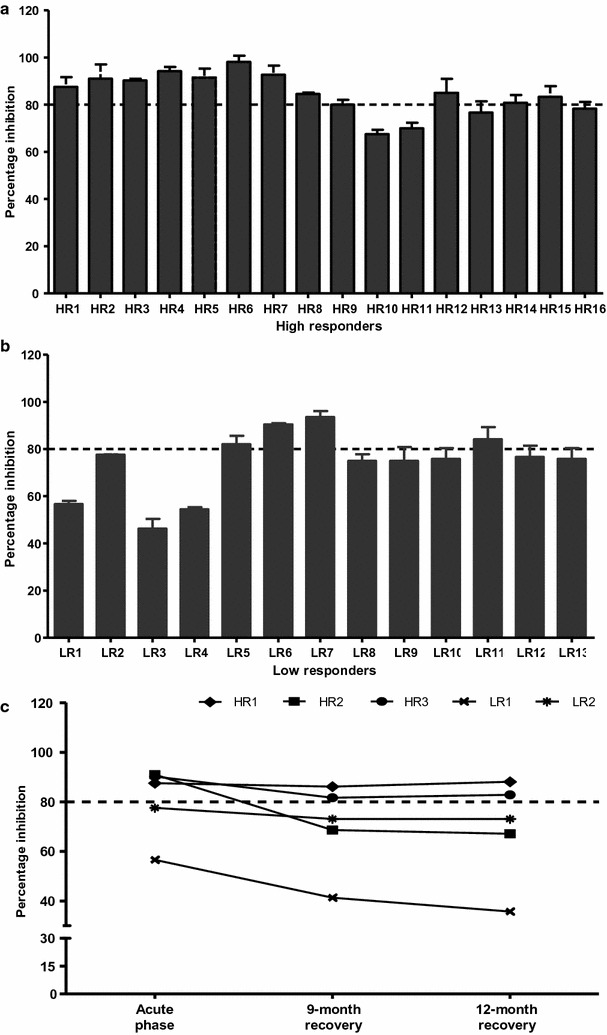
Methods: To evaluate the antibody responses to PvMSP1P-19 during and after recovery from P. vivax infection, heparinized blood was collected from P. vivax-infected patients and recovered subjects to detect the total IgG response. The seropositive samples were defined into high and low responders, according to their optical density (OD) values obtained from ELISA. High responders were the subjects who had OD values above the OD of antisera from non-exposed controls plus 4× standard deviations, whereas low responders were the subjects who had OD values less than OD of antisera from non-exposed controls plus 4× standard deviations. The plasma from high and low responders were taken for testing the inhibitory activity against PvMSP1P-19-erythrocyte binding by in vitro EBIA. The sustainability of PvMSP1P-19-specific memory B cell responses after recovery from infection was analysed by ELISPOT.
Results: The anti-PvMSP1P-19 antibody levels were significantly higher in acutely infected P. vivax patients compared to healthy controls (P < 0.0001). Monitoring of the anti-PvMSP1P-19 antibody titre showed that the antibody was maintained for up to 9 months after recovery. Almost all high-responder groups strongly inhibited PvMSP1P-19 binding to erythrocytes, whereas no inhibition was shown in most low-responder samples. Interestingly, the inhibitory activity of the antibodies in some individuals from high-responder samples were stable for at least 12 months. The longevity of the antibody response was associated with the presence of PvMSP1P-19-specific memory B cells at 9 months after recovery from infection.
Conclusions: The PvMSP1P-19 antigen has immunogenicity during the induction of the antibody response, in which both the levels and inhibitory activity are maintained after the patient recovered from P. vivax infection. The maintenance of the antibody response was associated with the response of PvMSP1P-19-specific memory B cells. Therefore, the PvMSP1P-19 antigen should also be considered as a reliable vaccine candidate to develop a blood-stage vaccine against P. vivax.
Keywords: Immunogenicity; Merozoite surface protein 1 paralog; Plasmodium vivax.
Figures



Similar articles
-
Development and longevity of naturally acquired antibody and memory B cell responses against Plasmodium vivax infection.PLoS Negl Trop Dis. 2024 Oct 24;18(10):e0012600. doi: 10.1371/journal.pntd.0012600. eCollection 2024 Oct. PLoS Negl Trop Dis. 2024. PMID: 39446698 Free PMC article. Review.
-
The Plasmodium vivax merozoite surface protein 1 paralog is a novel erythrocyte-binding ligand of P. vivax.Infect Immun. 2013 May;81(5):1585-95. doi: 10.1128/IAI.01117-12. Epub 2013 Mar 4. Infect Immun. 2013. PMID: 23460511 Free PMC article.
-
Naturally-acquired cellular immune response against Plasmodium vivax merozoite surface protein-1 paralog antigen.Malar J. 2015 Apr 15;14:159. doi: 10.1186/s12936-015-0681-8. Malar J. 2015. PMID: 25889175 Free PMC article.
-
The acquisition of long-lived memory B cell responses to merozoite surface protein-8 in individuals with Plasmodium vivax infection.Malar J. 2019 May 31;18(1):188. doi: 10.1186/s12936-019-2821-z. Malar J. 2019. PMID: 31151441 Free PMC article.
-
Vaccination for vivax malaria: targeting the invaders.Trends Parasitol. 2004 Mar;20(3):99-102. doi: 10.1016/j.pt.2003.12.003. Trends Parasitol. 2004. PMID: 16676415 Review.
Cited by
-
Elicitation of T-cell-derived IFN-γ-dependent immunity by highly conserved Plasmodium ovale curtisi Duffy binding protein domain region II (PocDBP-RII).Parasit Vectors. 2023 Aug 8;16(1):269. doi: 10.1186/s13071-023-05897-9. Parasit Vectors. 2023. PMID: 37553591 Free PMC article.
-
Navigating parasite antigen genetic diversity in the design of Plasmodium vivax serological exposure markers for malaria.bioRxiv [Preprint]. 2025 Jul 8:2025.07.07.663616. doi: 10.1101/2025.07.07.663616. bioRxiv. 2025. PMID: 40672192 Free PMC article. Preprint.
-
The persistence of naturally acquired antibodies and memory B cells specific to rhoptry proteins of Plasmodium vivax in patients from areas of low malaria transmission.Malar J. 2019 Nov 29;18(1):382. doi: 10.1186/s12936-019-3009-2. Malar J. 2019. PMID: 31783870 Free PMC article.
-
Development and longevity of naturally acquired antibody and memory B cell responses against Plasmodium vivax infection.PLoS Negl Trop Dis. 2024 Oct 24;18(10):e0012600. doi: 10.1371/journal.pntd.0012600. eCollection 2024 Oct. PLoS Negl Trop Dis. 2024. PMID: 39446698 Free PMC article. Review.
-
Development of B Cell Memory in Malaria.Front Immunol. 2019 Apr 2;10:559. doi: 10.3389/fimmu.2019.00559. eCollection 2019. Front Immunol. 2019. PMID: 31001244 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
