

The bioelectric code: An ancient computational medium for dynamic control of growth and form
- PMID: 28855098
- PMCID: PMC10464596
- DOI: 10.1016/j.biosystems.2017.08.009
The bioelectric code: An ancient computational medium for dynamic control of growth and form
Abstract
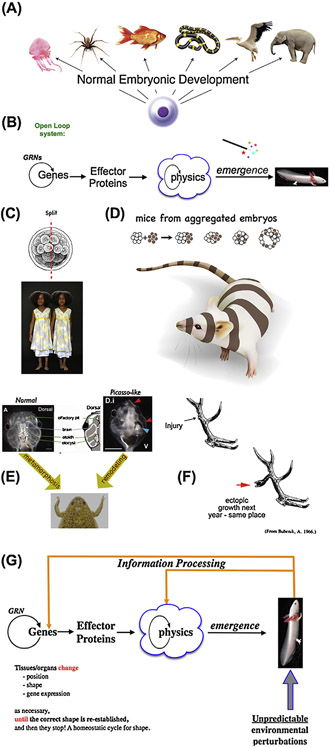
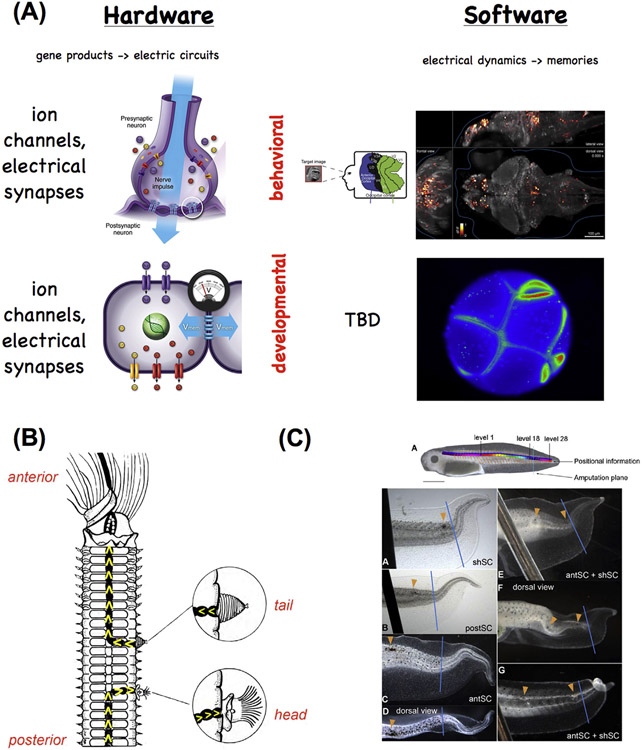
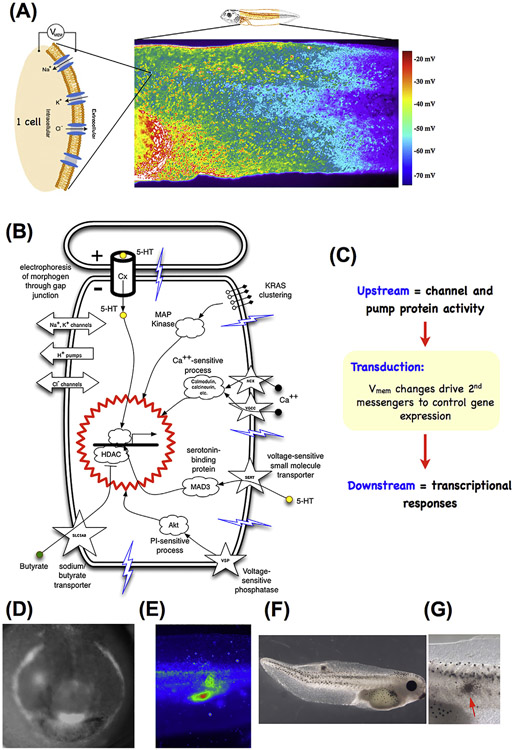
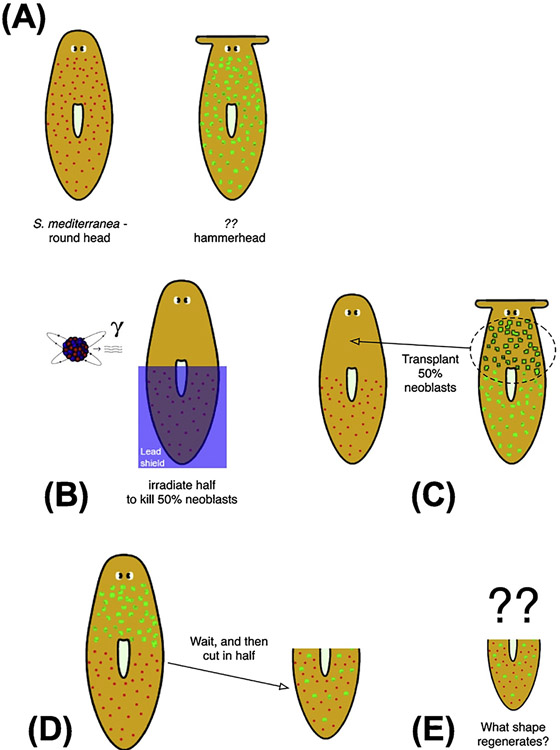
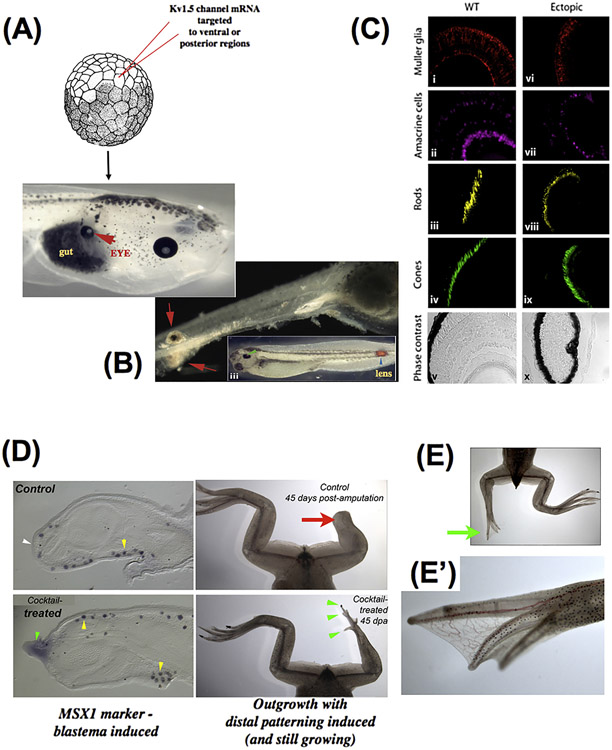
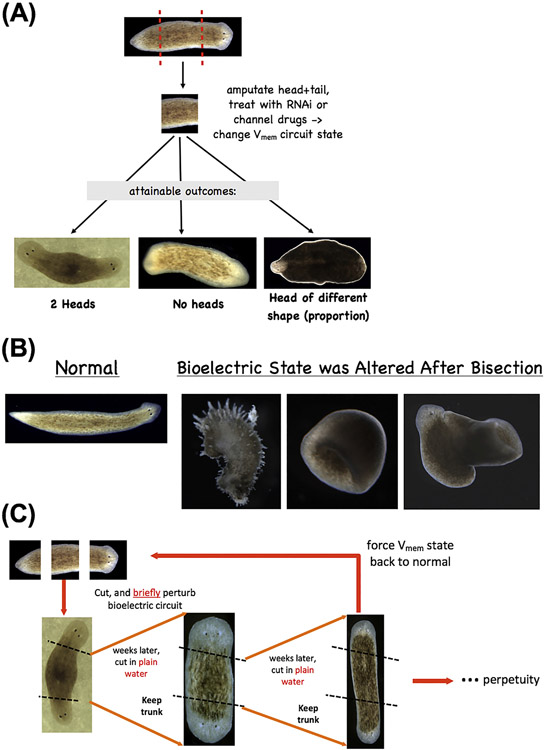
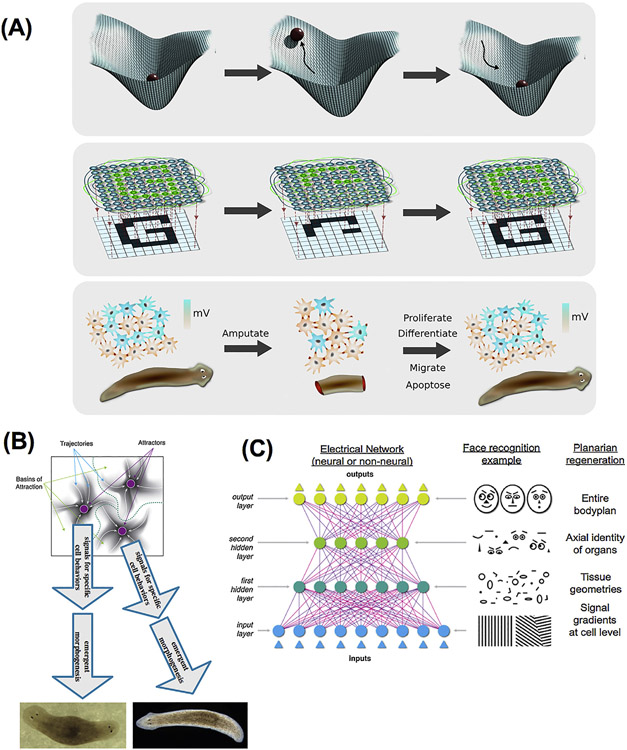
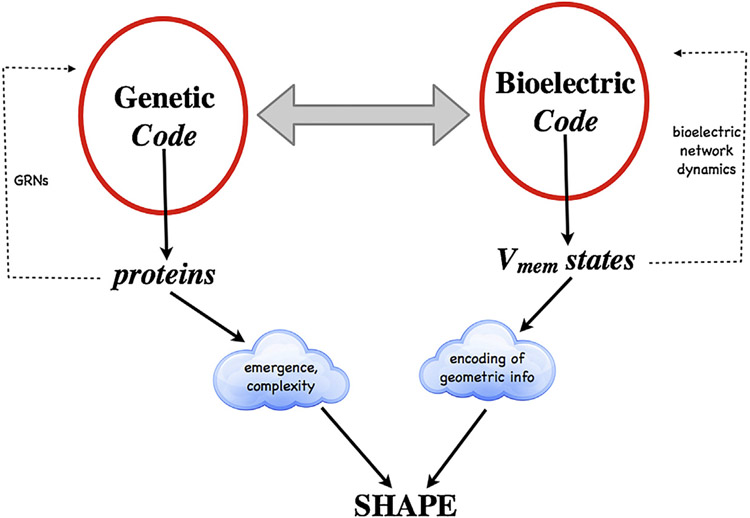
What determines large-scale anatomy? DNA does not directly specify geometrical arrangements of tissues and organs, and a process of encoding and decoding for morphogenesis is required. Moreover, many species can regenerate and remodel their structure despite drastic injury. The ability to obtain the correct target morphology from a diversity of initial conditions reveals that the morphogenetic code implements a rich system of pattern-homeostatic processes. Here, we describe an important mechanism by which cellular networks implement pattern regulation and plasticity: bioelectricity. All cells, not only nerves and muscles, produce and sense electrical signals; in vivo, these processes form bioelectric circuits that harness individual cell behaviors toward specific anatomical endpoints. We review emerging progress in reading and re-writing anatomical information encoded in bioelectrical states, and discuss the approaches to this problem from the perspectives of information theory, dynamical systems, and computational neuroscience. Cracking the bioelectric code will enable much-improved control over biological patterning, advancing basic evolutionary developmental biology as well as enabling numerous applications in regenerative medicine and synthetic bioengineering.
Keywords: Bayesian inference; Bioelectricity; Dynamical system theory; Embryogenesis; Ion channels; Morphogenesis; Patterning; Primitive cognition; Regeneration.
Copyright © 2017 Elsevier B.V. All rights reserved.
Figures
References
-
- Aboobaker AA, 2011. Planarian stem cells: a simple paradigm for regeneration. Trends Cell Biol. 21, 304–311. - PubMed
-
- Adams DS, Masi A, Levin M, 2007. H+ pump-dependent changes in membrane voltage are an early mechanism necessary and sufficient to induce Xenopus tail regeneration. Development 134, 1323–1335. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
