

Oligodendrocyte RasG12V expressed in its endogenous locus disrupts myelin structure through increased MAPK, nitric oxide, and notch signaling
- PMID: 28856719
- PMCID: PMC5766030
- DOI: 10.1002/glia.23209
Oligodendrocyte RasG12V expressed in its endogenous locus disrupts myelin structure through increased MAPK, nitric oxide, and notch signaling
Abstract
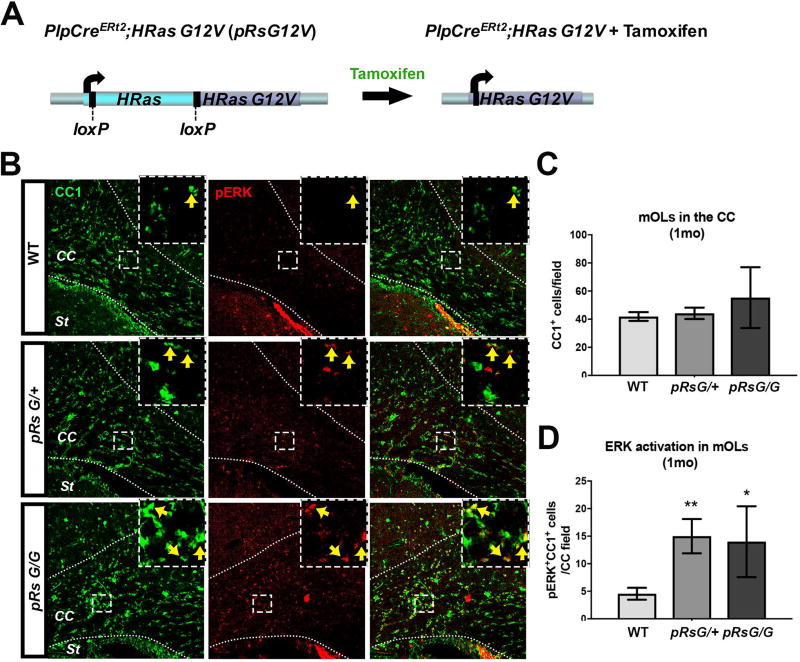
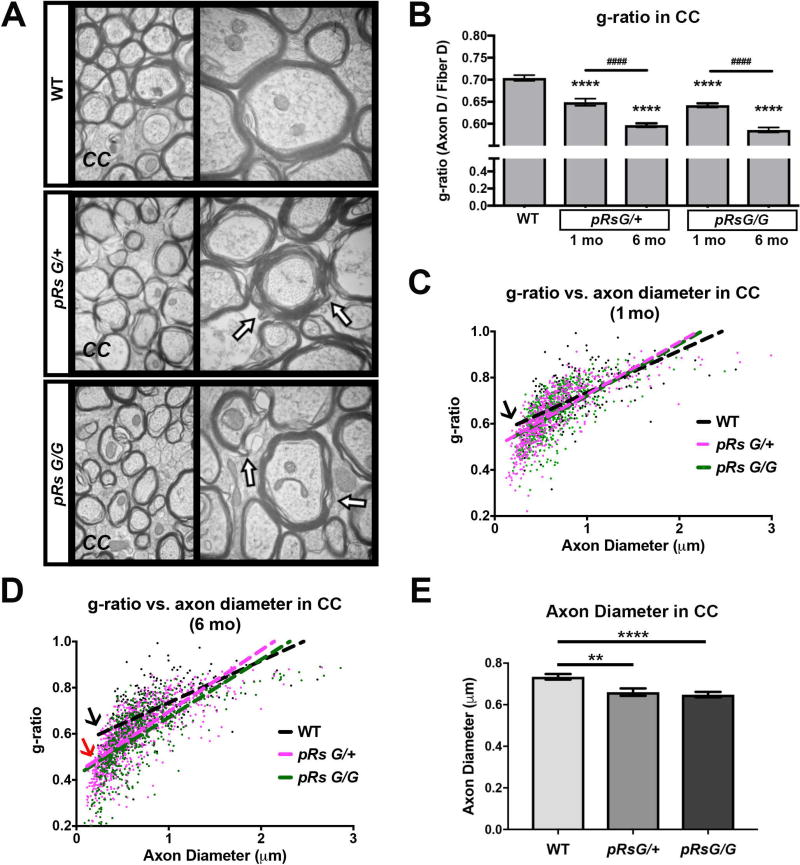
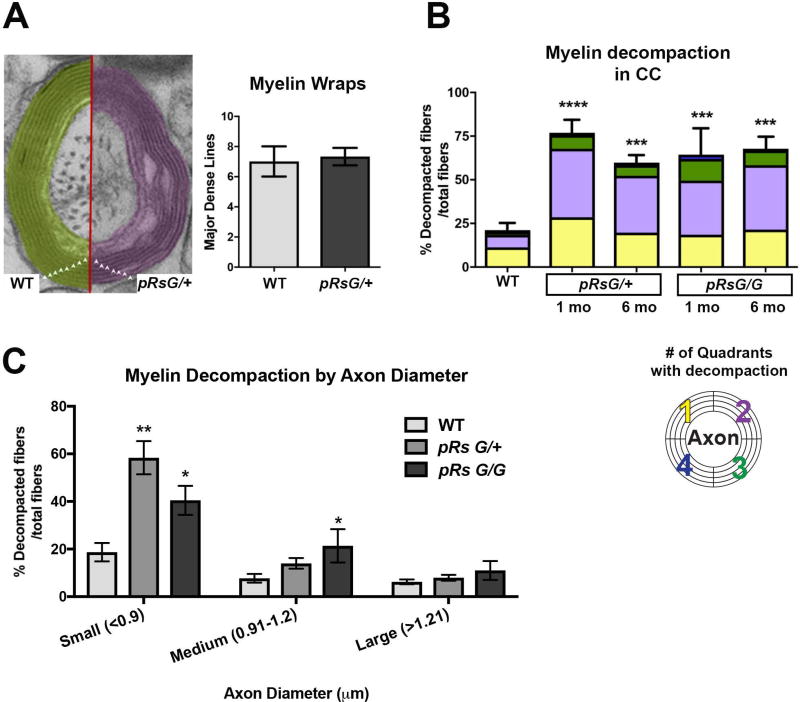
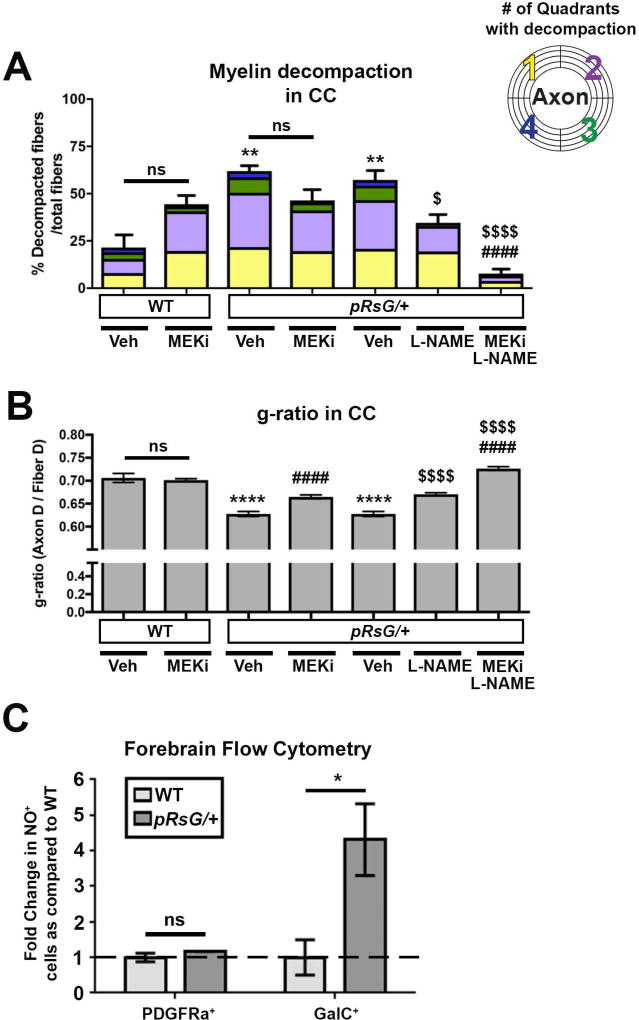
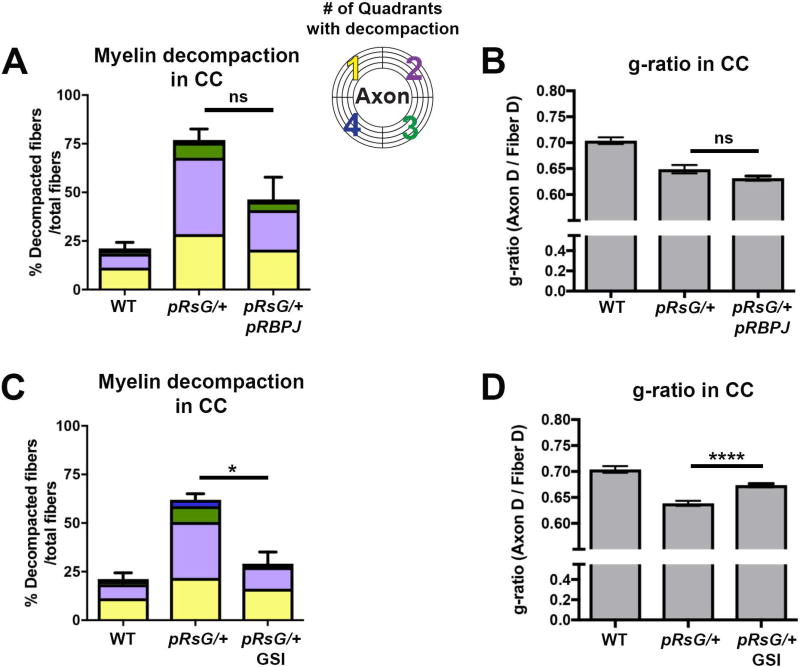
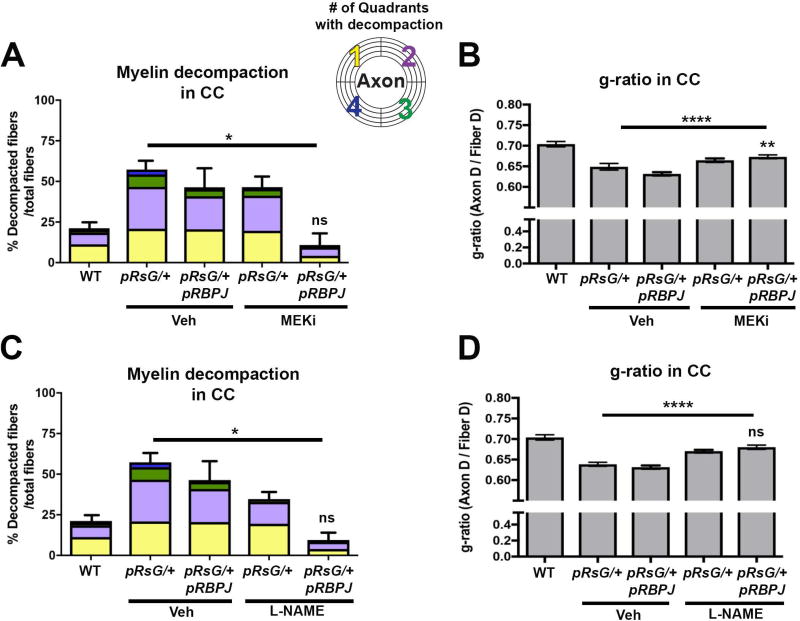
Costello syndrome (CS) is a gain of function Rasopathy caused by heterozygous activating mutations in the HRAS gene. Patients show brain dysfunction that can include abnormal brain white matter. Transgenic activation of HRas in the entire mouse oligodendrocyte lineage resulted in myelin defects and behavioral abnormalities, suggesting roles for disrupted myelin in CS brain dysfunction. Here, we studied a mouse model in which the endogenous HRas gene is conditionally replaced by mutant HRasG12V in mature oligodendrocytes, to separate effects in mature myelinating cells from developmental events. Increased myelin thickness due to decompaction was detectable within one month of HRasG12V expression in the corpus callosum of adult mice. Increases in active ERK and Nitric Oxide (NO) were present in HRas mutants and inhibition of NO synthase (NOS) or MEK each partially rescued myelin decompaction. In addition, genetic or pharmacologic inhibition of Notch signaling improved myelin compaction. Complete rescue of myelin structure required dual drug treatments combining MAPK, NO, or Notch inhibition; with MEK + NOS blockade producing the most robust effect. We suggest that individual or concomitant blockade of these pathways in CS patients may improve aspects of brain function.
Keywords: costello syndrome; myelin; nitric oxide; notch; oligodendrocyte; ras; rasopathy.
© 2017 Wiley Periodicals, Inc.
Figures
Similar articles
-
Oligodendrocyte Nf1 Controls Aberrant Notch Activation and Regulates Myelin Structure and Behavior.Cell Rep. 2017 Apr 18;19(3):545-557. doi: 10.1016/j.celrep.2017.03.073. Cell Rep. 2017. PMID: 28423318 Free PMC article.
-
Strength of ERK1/2 MAPK Activation Determines Its Effect on Myelin and Axonal Integrity in the Adult CNS.J Neurosci. 2016 Jun 15;36(24):6471-87. doi: 10.1523/JNEUROSCI.0299-16.2016. J Neurosci. 2016. PMID: 27307235 Free PMC article.
-
Nf1 loss and Ras hyperactivation in oligodendrocytes induce NOS-driven defects in myelin and vasculature.Cell Rep. 2013 Sep 26;4(6):1197-212. doi: 10.1016/j.celrep.2013.08.011. Epub 2013 Sep 12. Cell Rep. 2013. PMID: 24035394 Free PMC article.
-
ERK1/2 Activation in Preexisting Oligodendrocytes of Adult Mice Drives New Myelin Synthesis and Enhanced CNS Function.J Neurosci. 2016 Aug 31;36(35):9186-200. doi: 10.1523/JNEUROSCI.1444-16.2016. J Neurosci. 2016. PMID: 27581459 Free PMC article.
-
Cross-talk between F3/contactin and Notch at axoglial interface: a role in oligodendrocyte development.Dev Neurosci. 2006;28(1-2):25-33. doi: 10.1159/000090750. Dev Neurosci. 2006. PMID: 16508301 Review.
Cited by
-
Daam2 Regulates Myelin Structure and the Oligodendrocyte Actin Cytoskeleton through Rac1 and Gelsolin.J Neurosci. 2022 Mar 2;42(9):1679-1691. doi: 10.1523/JNEUROSCI.1517-21.2022. Epub 2022 Jan 31. J Neurosci. 2022. PMID: 35101966 Free PMC article.
-
Protection of Oligodendrocytes Through Neuronal Overexpression of the Small GTPase Ras in Hyperoxia-Induced Neonatal Brain Injury.Front Neurol. 2018 Mar 21;9:175. doi: 10.3389/fneur.2018.00175. eCollection 2018. Front Neurol. 2018. PMID: 29619004 Free PMC article.
-
Three dimensional electron microscopy reveals changing axonal and myelin morphology along normal and partially injured optic nerves.Sci Rep. 2018 Mar 5;8(1):3979. doi: 10.1038/s41598-018-22361-2. Sci Rep. 2018. PMID: 29507421 Free PMC article.
-
Generation of a Mouse Model to Study the Noonan Syndrome Gene Lztr1 in the Telencephalon.Front Cell Dev Biol. 2021 Jun 16;9:673995. doi: 10.3389/fcell.2021.673995. eCollection 2021. Front Cell Dev Biol. 2021. PMID: 34222248 Free PMC article.
-
The sixth international RASopathies symposium: Precision medicine-From promise to practice.Am J Med Genet A. 2020 Mar;182(3):597-606. doi: 10.1002/ajmg.a.61434. Epub 2019 Dec 11. Am J Med Genet A. 2020. PMID: 31825160 Free PMC article.
References
-
- Aoki Y, Niihori T, Kawame H, Kurosawa K, Ohashi H, Tanaka Y, Filocamo M, Kato K, Suzuki Y, Kure S, et al. Germline mutations in HRAS proto-oncogene cause Costello syndrome. Nat Genet. 2005;37:1038–40. - PubMed
-
- Axelrad ME, Nicholson L, Stabley DL, Sol-Church K, Gripp KW. Longitudinal assessment of cognitive characteristics in Costello syndrome. Am J Med Genet A. 2007;143A:3185–93. - PubMed
-
- Barrett SD, Bridges AJ, Dudley DT, Saltiel AR, Fergus JH, Flamme CM, Delaney AM, Kaufman M, LePage S, Leopold WR, et al. The discovery of the benzhydroxamate MEK inhibitors CI-1040 and PD 0325901. Bioorg Med Chem Lett. 2008;18:6501–4. - PubMed
-
- Bauer NG, Richter-Landsberg C, Ffrench-Constant C. Role of the oligodendroglial cytoskeleton in differentiation and myelination. Glia. 2009;57:1691–705. - PubMed
-
- Baumgart A, Mazur PK, Anton M, Rudelius M, Schwamborn K, Feuchtinger A, Behnke K, Walch A, Braren R, Peschel C, et al. Opposing role of Notch1 and Notch2 in a Kras(G12D)-driven murine non-small cell lung cancer model. Oncogene. 2015;34:578–88. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
