

CCCTC-binding factor is essential to the maintenance and quiescence of hematopoietic stem cells in mice
- PMID: 28857086
- PMCID: PMC5579513
- DOI: 10.1038/emm.2017.124
CCCTC-binding factor is essential to the maintenance and quiescence of hematopoietic stem cells in mice
Abstract
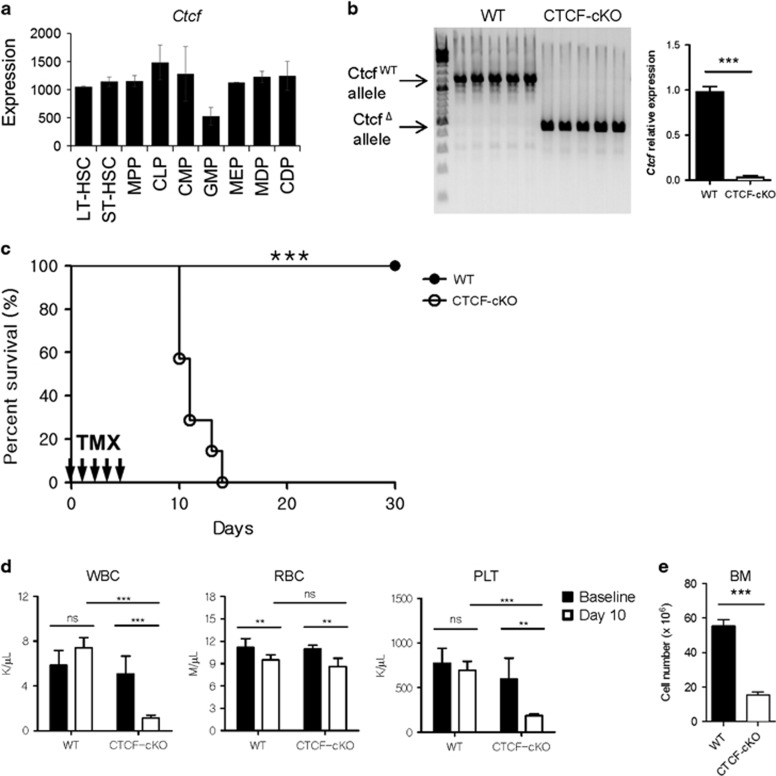
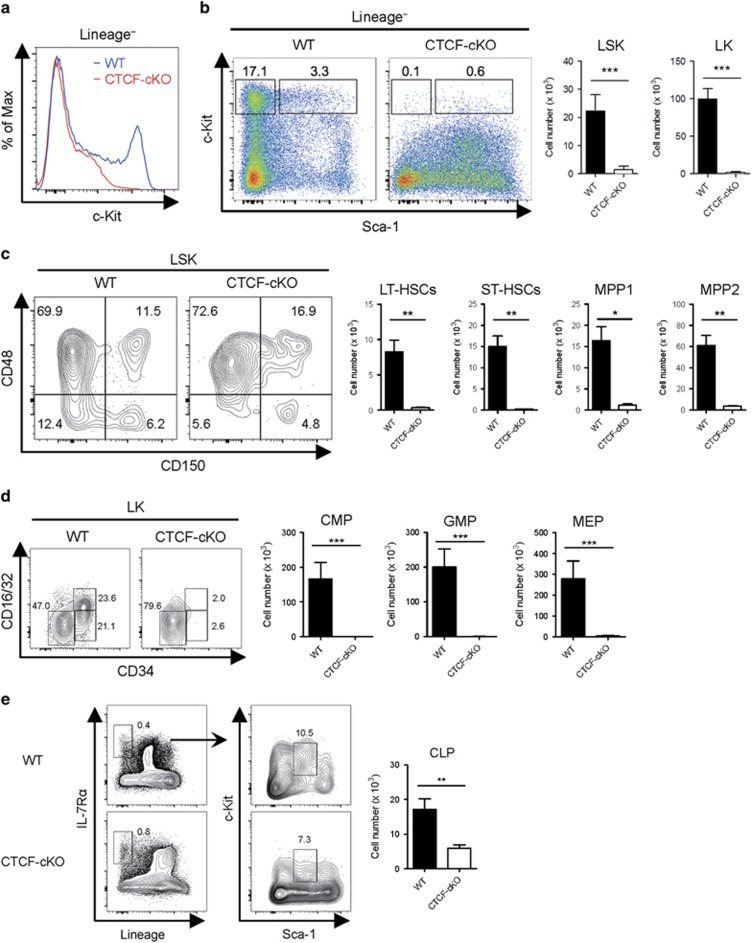
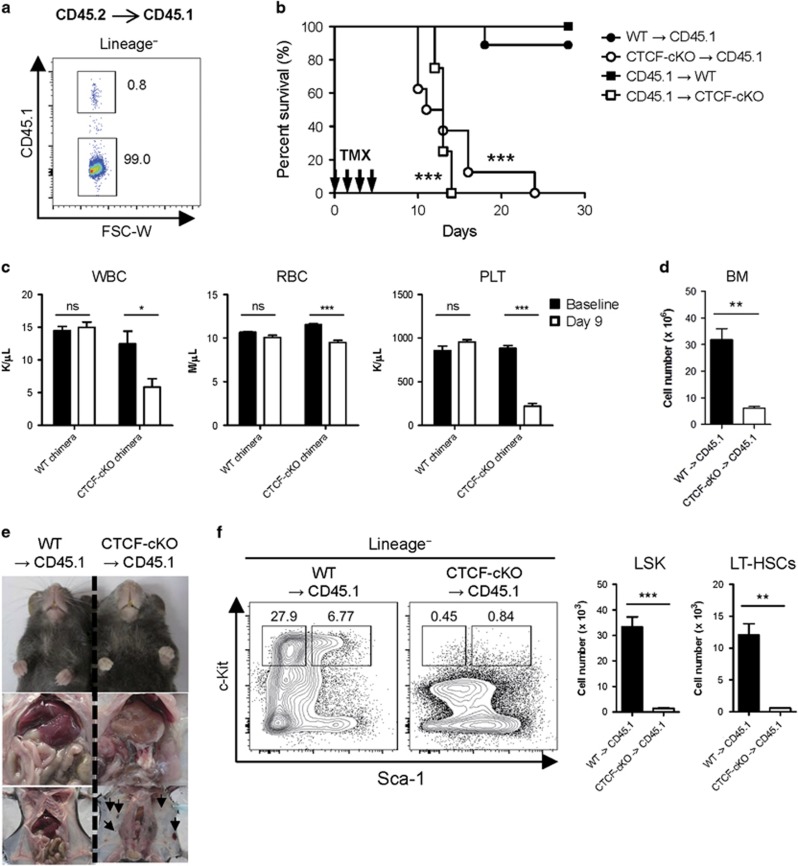
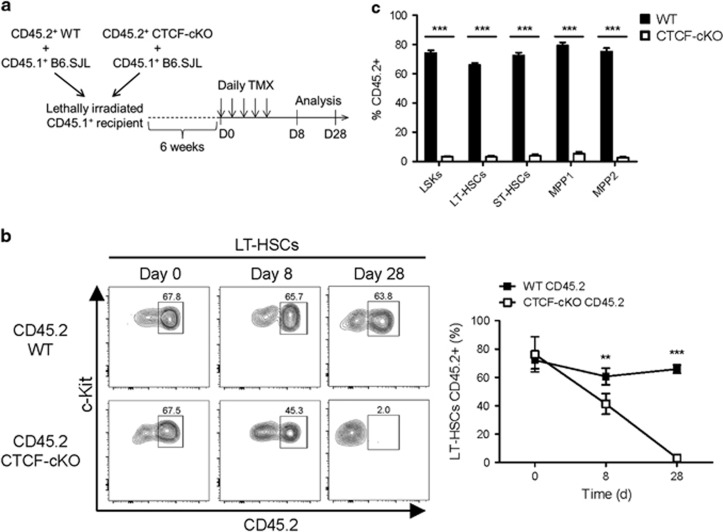
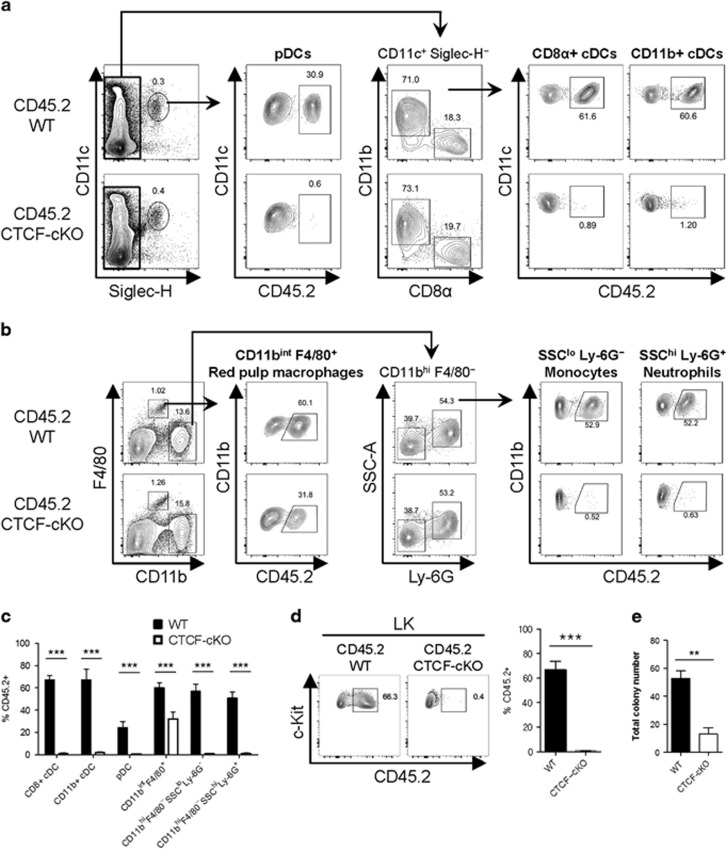
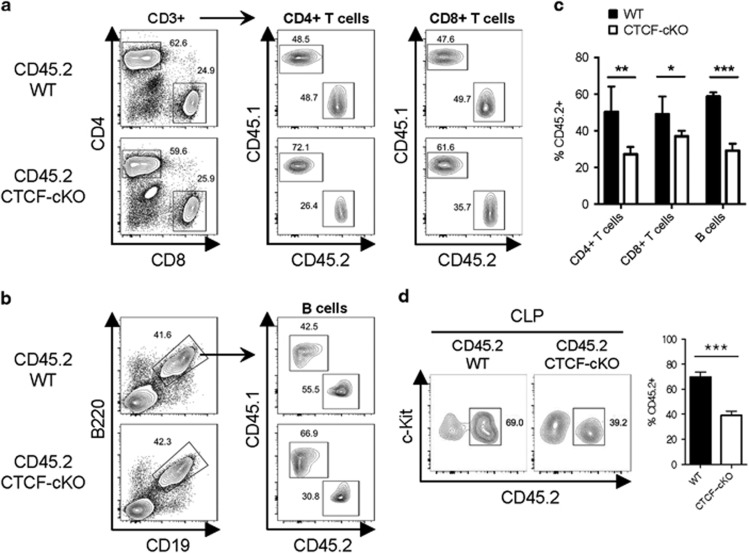
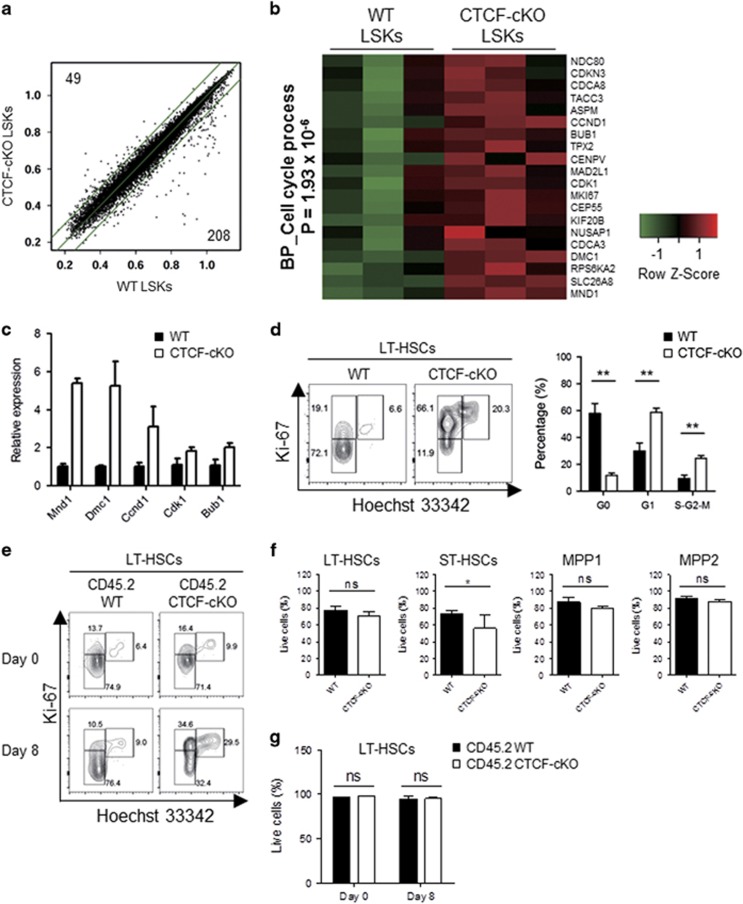
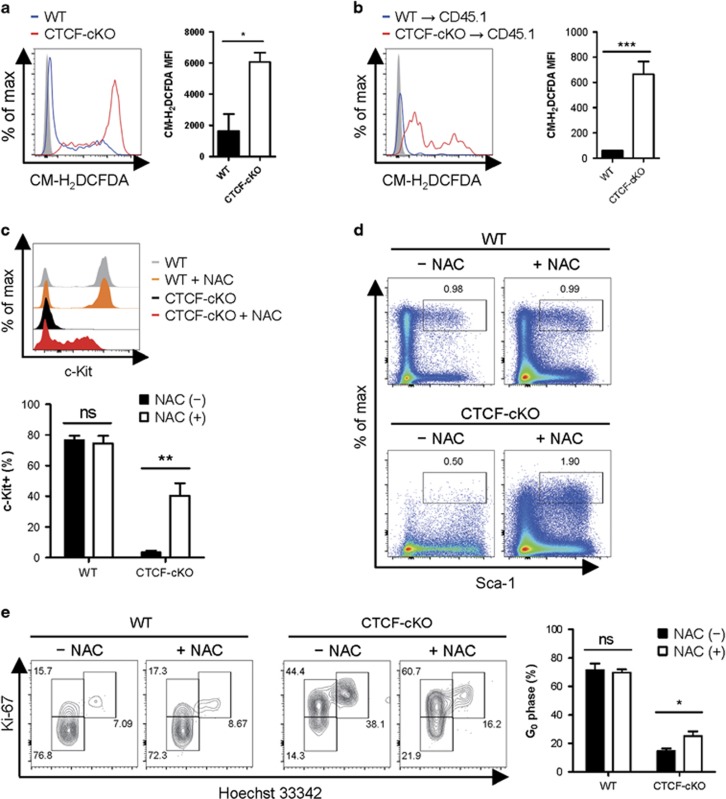
Hematopoiesis involves a series of lineage differentiation programs initiated in hematopoietic stem cells (HSCs) found in bone marrow (BM). To ensure lifelong hematopoiesis, various molecular mechanisms are needed to maintain the HSC pool. CCCTC-binding factor (CTCF) is a DNA-binding, zinc-finger protein that regulates the expression of its target gene by organizing higher order chromatin structures. Currently, the role of CTCF in controlling HSC homeostasis is unknown. Using a tamoxifen-inducible CTCF conditional knockout mouse system, we aimed to determine whether CTCF regulates the homeostatic maintenance of HSCs. In adult mice, acute systemic CTCF ablation led to severe BM failure and the rapid shrinkage of multiple c-Kithi progenitor populations, including Sca-1+ HSCs. Similarly, hematopoietic system-confined CTCF depletion caused an acute loss of HSCs and highly increased mortality. Mixed BM chimeras reconstituted with supporting BM demonstrated that CTCF deficiency-mediated HSC depletion has both cell-extrinsic and cell-intrinsic effects. Although c-Kithi myeloid progenitor cell populations were severely reduced after ablating Ctcf, c-Kitint common lymphoid progenitors and their progenies were less affected by the lack of CTCF. Whole-transcriptome microarray and cell cycle analyses indicated that CTCF deficiency results in the enhanced expression of the cell cycle-promoting program, and that CTCF-depleted HSCs express higher levels of reactive oxygen species (ROS). Importantly, in vivo treatment with an antioxidant partially rescued c-Kithi cell populations and their quiescence. Altogether, our results suggest that CTCF is indispensable for maintaining adult HSC pools, likely by regulating ROS-dependent HSC quiescence.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Wilson A, Trumpp A. Bone-marrow haematopoietic-stem-cell niches. Nat Rev Immunol 2006; 6: 93–106. - PubMed
-
- Wilson NK, Foster SD, Wang X, Knezevic K, Schutte J, Kaimakis P et al. Combinatorial transcriptional control in blood stem/progenitor cells: genome-wide analysis of ten major transcriptional regulators. Cell Stem Cell 2010; 7: 532–544. - PubMed
-
- Bigas A, Espinosa L. Hematopoietic stem cells: to be or Notch to be. Blood 2012; 119: 3226–3235. - PubMed
-
- Bigas A, Guiu J, Gama-Norton L. Notch and Wnt signaling in the emergence of hematopoietic stem cells. Blood Cells Mol Dis 2013; 51: 264–270. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
