

Principles of cellular resource allocation revealed by condition-dependent proteome profiling
- PMID: 28857745
- PMCID: PMC5578734
- DOI: 10.7554/eLife.28034
Principles of cellular resource allocation revealed by condition-dependent proteome profiling
Abstract
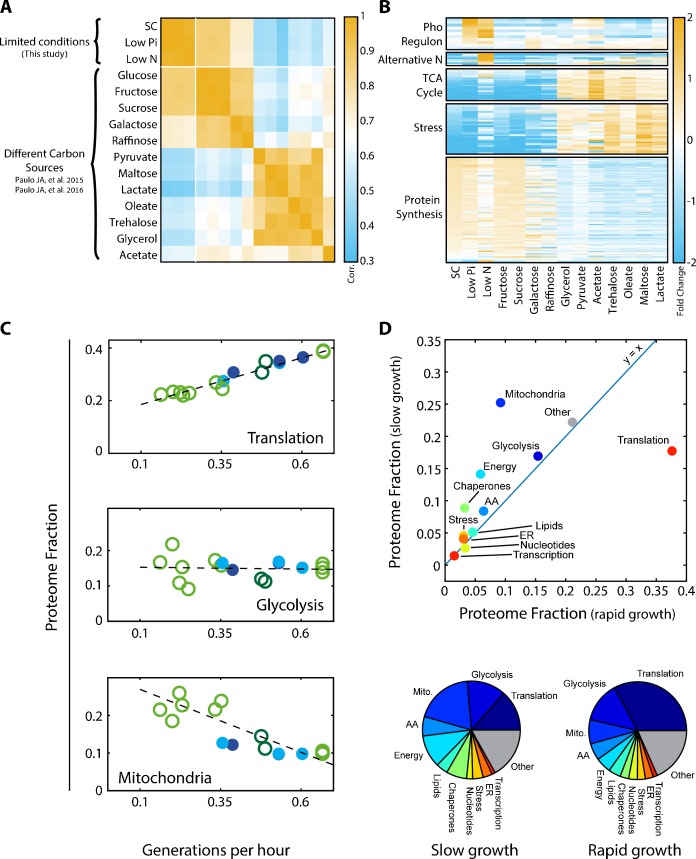
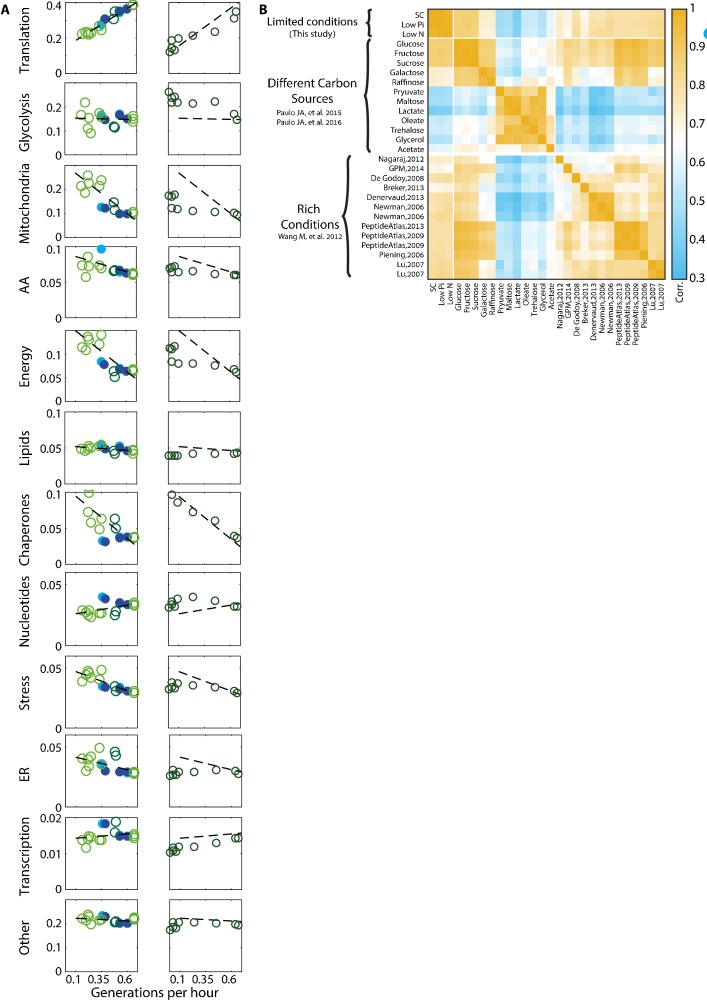
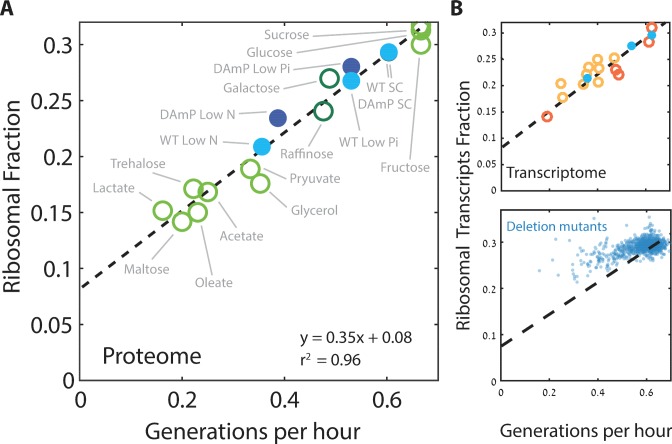
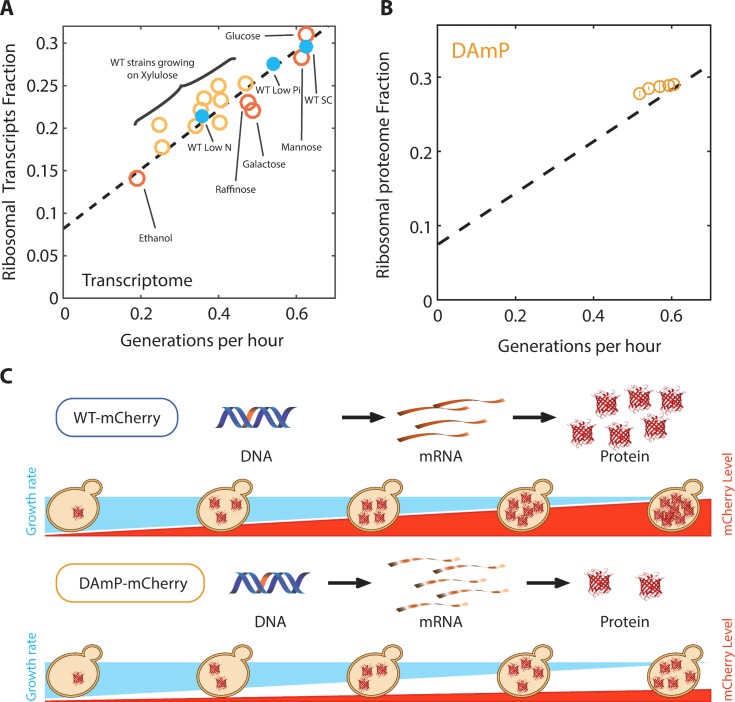
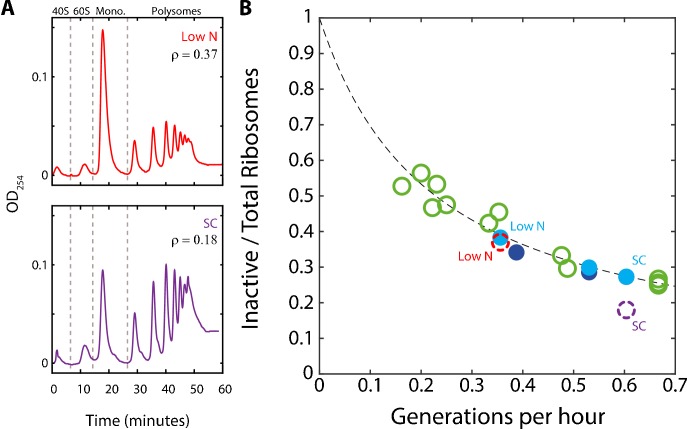
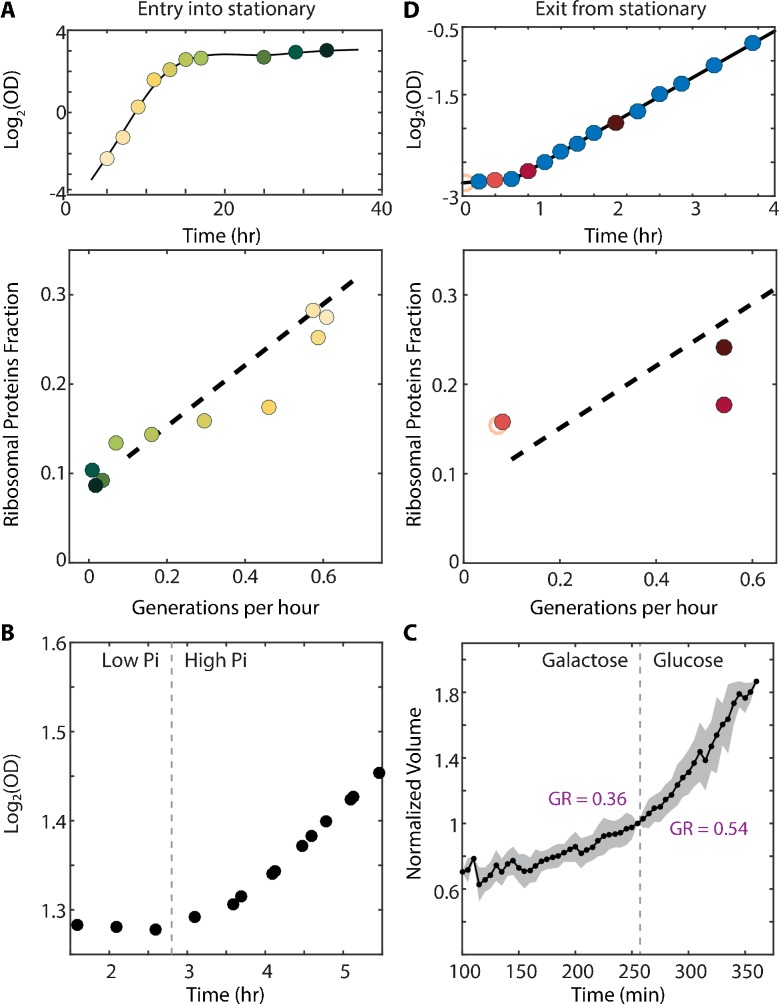
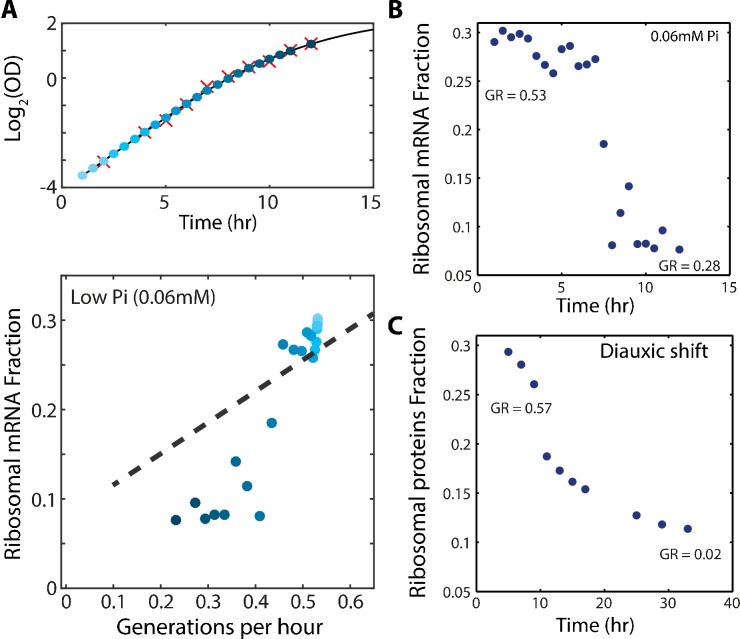
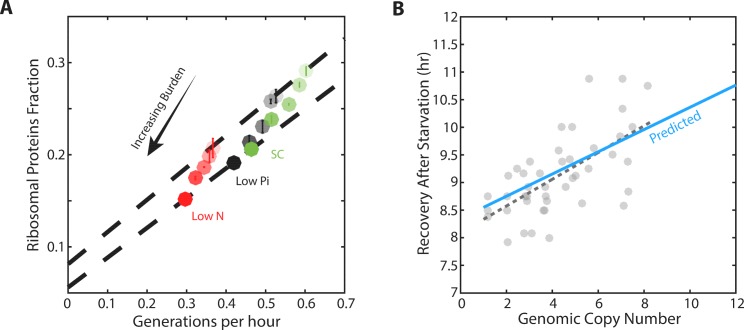
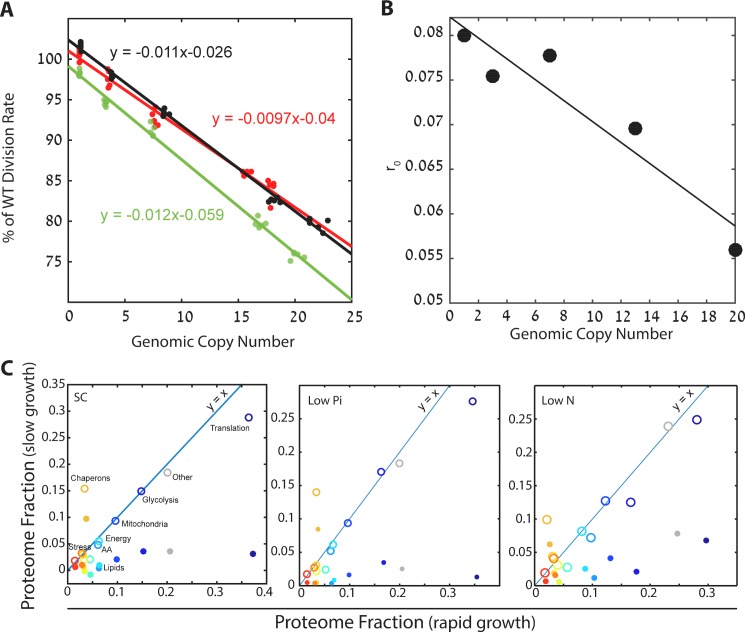
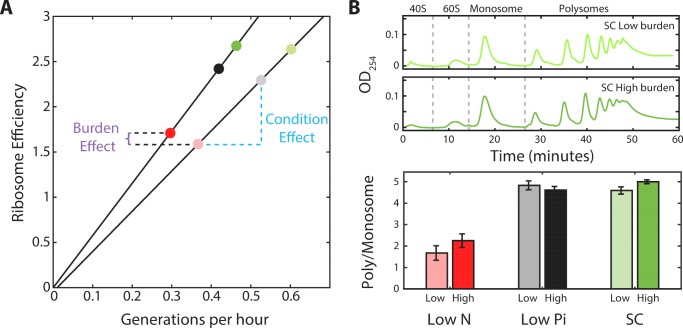
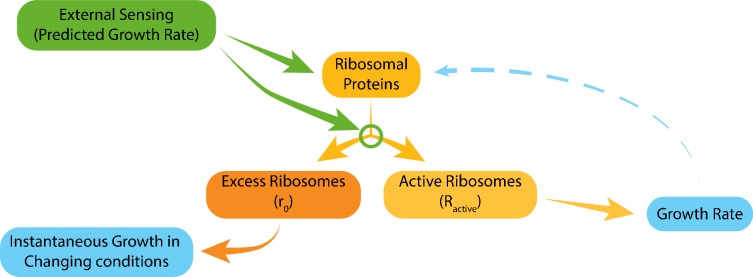
Growing cells coordinate protein translation with metabolic rates. Central to this coordination is ribosome production. Ribosomes drive cell growth, but translation of ribosomal proteins competes with production of non-ribosomal proteins. Theory shows that cell growth is maximized when all expressed ribosomes are constantly translating. To examine whether budding yeast function at this limit of full ribosomal usage, we profiled the proteomes of cells growing in different environments. We find that cells produce excess ribosomal proteins, amounting to a constant ≈8% of the proteome. Accordingly, ≈25% of ribosomal proteins expressed in rapidly growing cells does not contribute to translation. Further, this fraction increases as growth rate decreases and these excess ribosomal proteins are employed when translation demands unexpectedly increase. We suggest that steadily growing cells prepare for conditions that demand increased translation by producing excess ribosomes, at the expense of lower steady-state growth rate.
Keywords: S. cerevisiae; cell biology; computational biology; proteome; resource allocation; ribosomes; systems biology; transcription; translation.
Conflict of interest statement
Senior editor, eLife.
No competing interests declared.
Figures
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
