

Optimized retroviral transduction of mouse T cells for in vivo assessment of gene function
- PMID: 28858287
- PMCID: PMC6020692
- DOI: 10.1038/nprot.2017.083
Optimized retroviral transduction of mouse T cells for in vivo assessment of gene function
Abstract
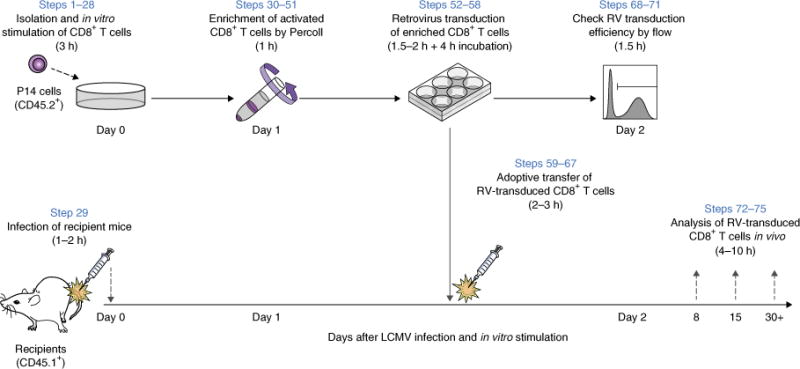
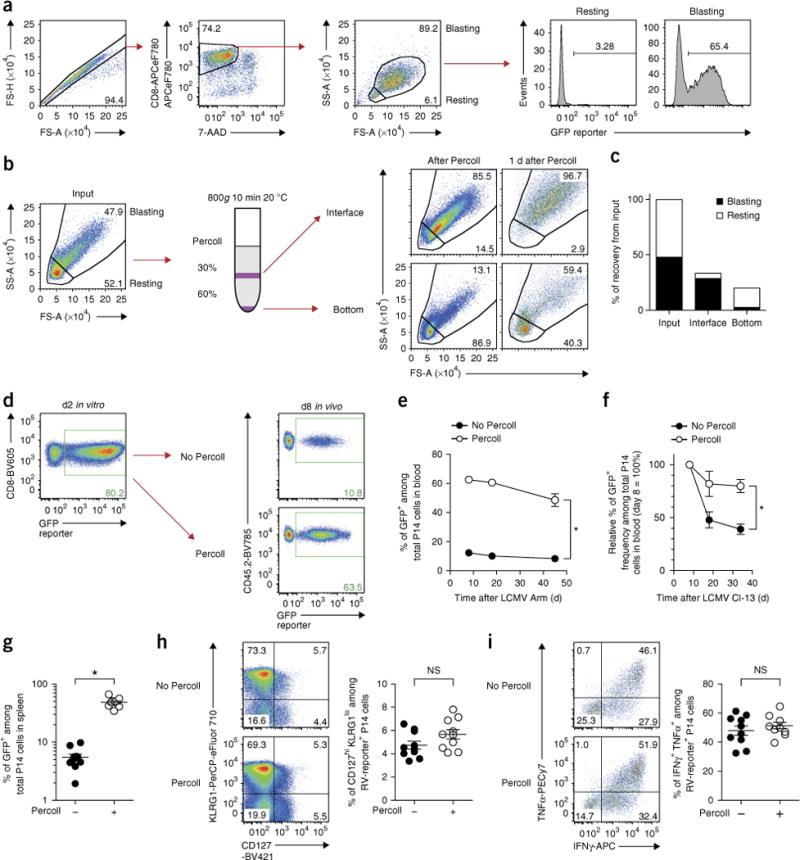
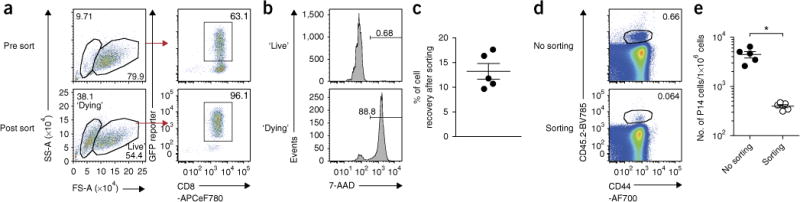
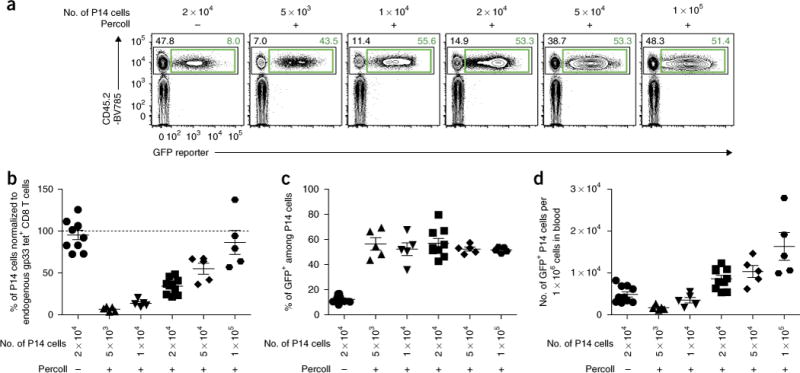
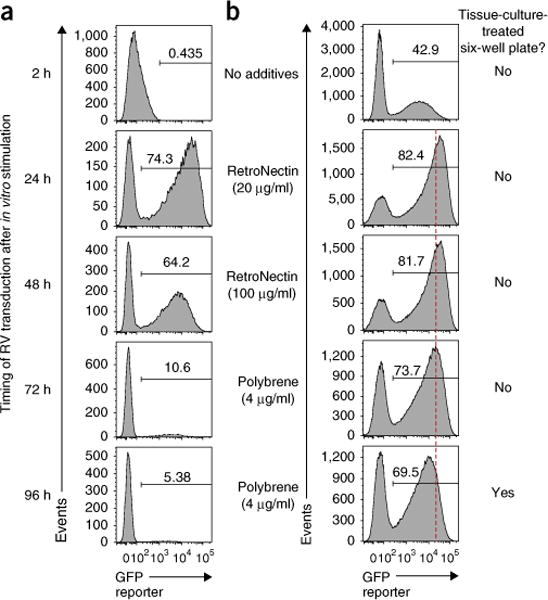
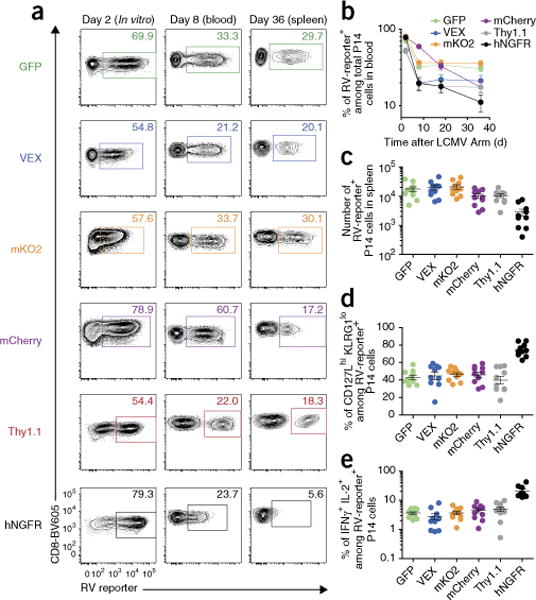
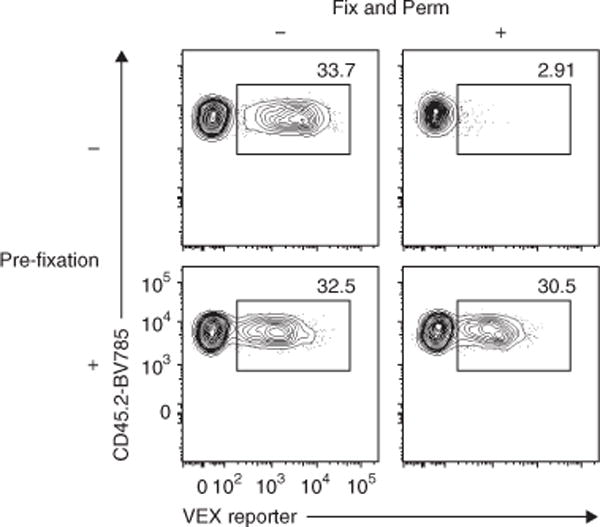
Retroviral (RV) expression of genes of interest (GOIs) is an invaluable tool and has formed the foundation of cellular engineering for adoptive cell therapy in cancer and other diseases. However, monitoring of transduced T cells long term (weeks to months) in vivo remains challenging because of the low frequency and often poor durability of transduced T cells over time when transferred without enrichment. Traditional methods often require additional overnight in vitro culture after transduction. Moreover, in vitro-generated effector CD8+ T cells enriched by sorting often have reduced viability, making it difficult to monitor the fate of transferred cells in vivo. Here, we describe an optimized mouse CD8+ T-cell RV transduction protocol that uses simple and rapid Percoll density centrifugation to enrich RV-susceptible activated CD8+ T cells. Percoll density centrifugation is simple, can be done on the day of transduction, requires minimal time, has low reagent costs and improves cell recovery (up to 60%), as well as the frequency of RV-transduced cells (∼sixfold over several weeks in vivo as compared with traditional methods). We have used this protocol to assess the long-term stability of CD8+ T cells after RV transduction by comparing the durability of T cells transduced with retroviruses expressing each of six commonly used RV reporter genes. Thus, we provide an optimized enrichment and transduction approach that allows long-term in vivo assessment of RV-transduced T cells. The overall procedure from T-cell isolation to RV transduction takes 2 d, and enrichment of activated T cells can be done in 1 h.
Conflict of interest statement
Figures
References
-
- Masopust D, Schenkel JM. The integration of T cell migration, differentiation and function. Nat Rev Immunol. 2013;13:309–320. - PubMed
-
- Cepko C, Pear W. Overview of the retrovirus transduction system. Curr Protoc Mol Biol. 2001 Chapter 9, Unit 9 9. - PubMed
-
- Lee J, Sadelain M, Brentjens R. In: Genetic Modification of Hematopoietic Stem Cells: Methods and Protocols. Baum C, editor. Humana Press; 2009. pp. 83–96.
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
