

Exercise-Induced Changes in Glucose Metabolism Promote Physiological Cardiac Growth
- PMID: 28860122
- PMCID: PMC5704654
- DOI: 10.1161/CIRCULATIONAHA.117.028274
Exercise-Induced Changes in Glucose Metabolism Promote Physiological Cardiac Growth
Abstract
Background: Exercise promotes metabolic remodeling in the heart, which is associated with physiological cardiac growth; however, it is not known whether or how physical activity-induced changes in cardiac metabolism cause myocardial remodeling. In this study, we tested whether exercise-mediated changes in cardiomyocyte glucose metabolism are important for physiological cardiac growth.
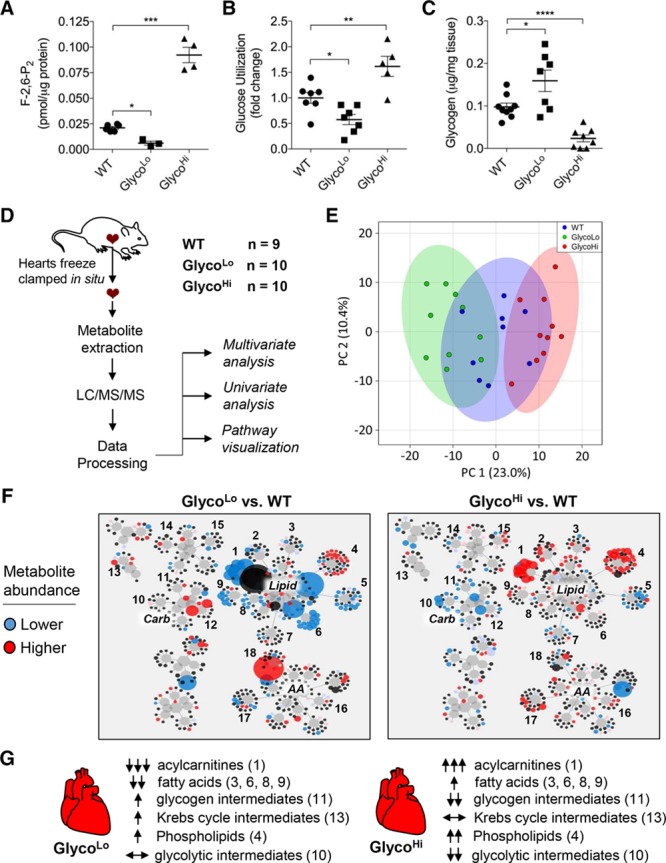
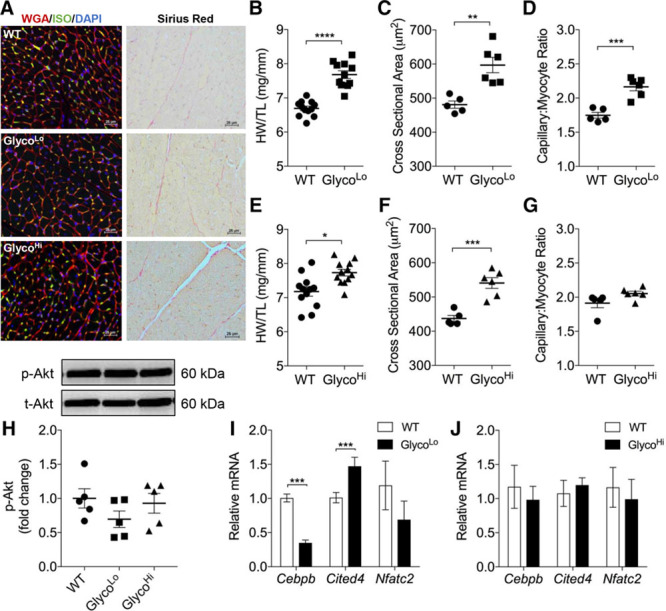
Methods: We used radiometric, immunologic, metabolomic, and biochemical assays to measure changes in myocardial glucose metabolism in mice subjected to acute and chronic treadmill exercise. To assess the relevance of changes in glycolytic activity, we determined how cardiac-specific expression of mutant forms of 6-phosphofructo-2-kinase/fructose-2,6-bisphosphatase affect cardiac structure, function, metabolism, and gene programs relevant to cardiac remodeling. Metabolomic and transcriptomic screenings were used to identify metabolic pathways and gene sets regulated by glycolytic activity in the heart.
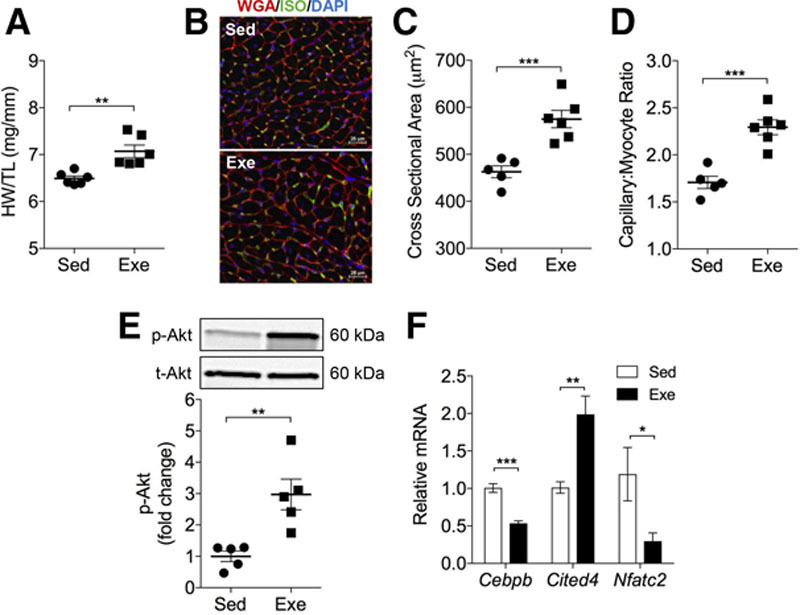
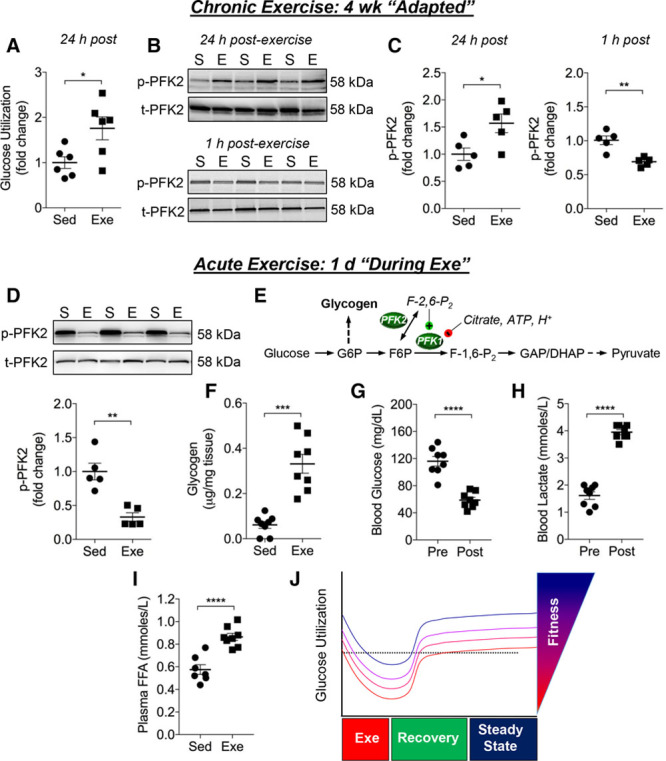
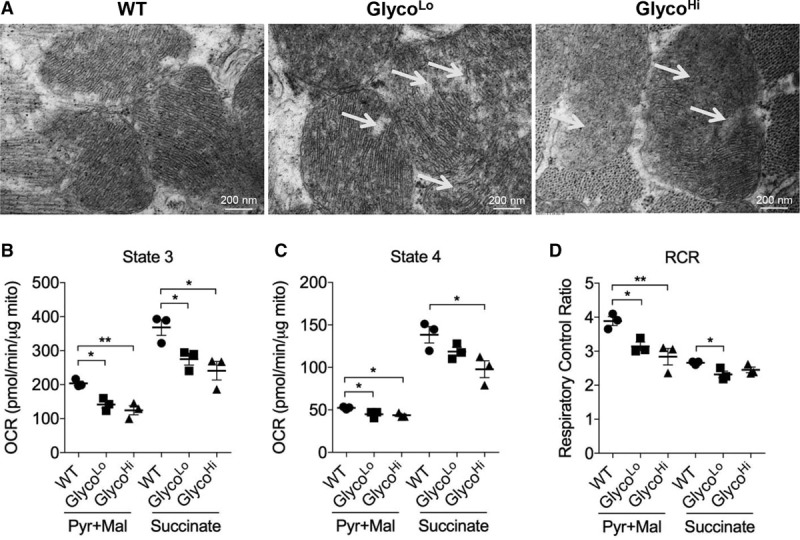
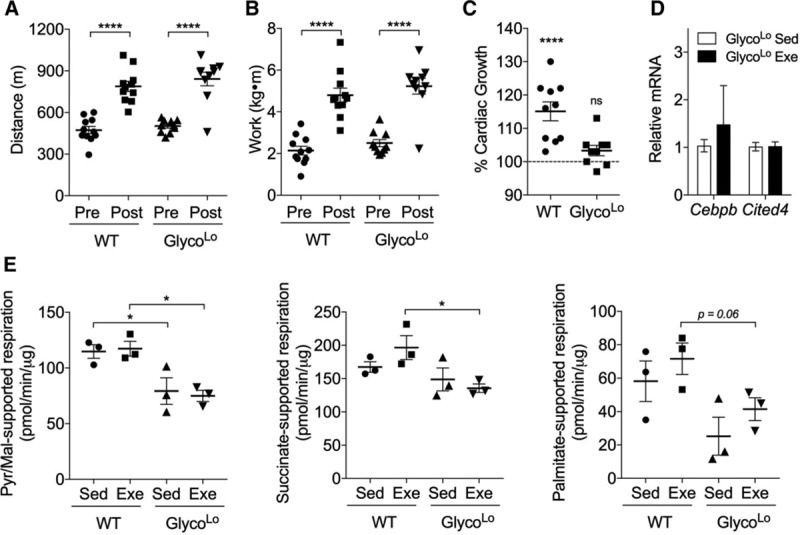
Results: Exercise acutely decreased glucose utilization via glycolysis by modulating circulating substrates and reducing phosphofructokinase activity; however, in the recovered state following exercise adaptation, there was an increase in myocardial phosphofructokinase activity and glycolysis. In mice, cardiac-specific expression of a kinase-deficient 6-phosphofructo-2-kinase/fructose-2,6-bisphosphatase transgene (GlycoLo mice) lowered glycolytic rate and regulated the expression of genes known to promote cardiac growth. Hearts of GlycoLo mice had larger myocytes, enhanced cardiac function, and higher capillary-to-myocyte ratios. Expression of phosphatase-deficient 6-phosphofructo-2-kinase/fructose-2,6-bisphosphatase in the heart (GlycoHi mice) increased glucose utilization and promoted a more pathological form of hypertrophy devoid of transcriptional activation of the physiological cardiac growth program. Modulation of phosphofructokinase activity was sufficient to regulate the glucose-fatty acid cycle in the heart; however, metabolic inflexibility caused by invariantly low or high phosphofructokinase activity caused modest mitochondrial damage. Transcriptomic analyses showed that glycolysis regulates the expression of key genes involved in cardiac metabolism and remodeling.
Conclusions: Exercise-induced decreases in glycolytic activity stimulate physiological cardiac remodeling, and metabolic flexibility is important for maintaining mitochondrial health in the heart.
Keywords: exercise; glycolysis; hypertrophy; metabolomics; mitochondria; ventricular remodeling.
© 2017 The Authors.
Figures
Comment in
-
Metabolism: A Direct Link Between Cardiac Structure and Function.Circulation. 2017 Nov 28;136(22):2158-2161. doi: 10.1161/CIRCULATIONAHA.117.031372. Circulation. 2017. PMID: 29180494 Free PMC article. No abstract available.
References
-
- Egan B, Zierath JR. Exercise metabolism and the molecular regulation of skeletal muscle adaptation. Cell Metab. 2013;17:162–184. doi: 10.1016/j.cmet.2012.12.012. - PubMed
-
- López-Otín C, Galluzzi L, Freije JM, Madeo F, Kroemer G. Metabolic control of longevity. Cell. 2016;166:802–821. doi: 10.1016/j.cell.2016.07.031. - PubMed
-
- Bassett DR, Jr, Howley ET. Limiting factors for maximum oxygen uptake and determinants of endurance performance. Med Sci Sports Exerc. 2000;32:70–84. - PubMed
-
- Wilson MG, Ellison GM, Cable NT. Basic science behind the cardiovascular benefits of exercise. Br J Sports Med. 2016;50:93–99. doi: 10.1136/bjsports-2014-306596rep. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
