

Discovery of a highly selective chemical inhibitor of matrix metalloproteinase-9 (MMP-9) that allosterically inhibits zymogen activation
- PMID: 28860188
- PMCID: PMC5663893
- DOI: 10.1074/jbc.M117.806075
Discovery of a highly selective chemical inhibitor of matrix metalloproteinase-9 (MMP-9) that allosterically inhibits zymogen activation
Abstract
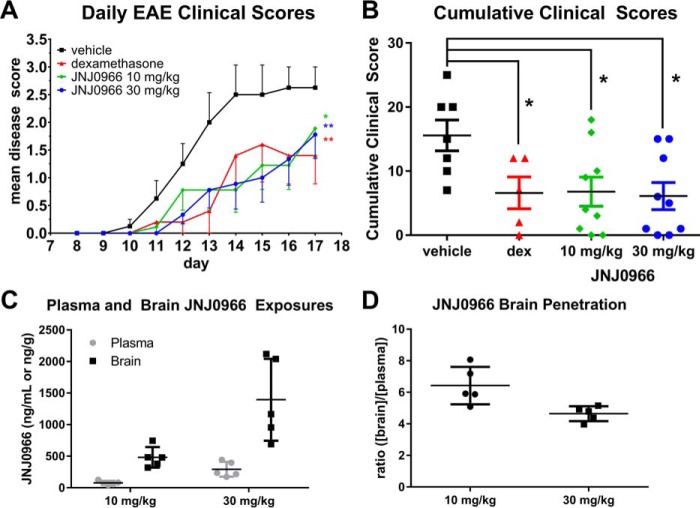
Aberrant activation of matrix metalloproteinases (MMPs) is a common feature of pathological cascades observed in diverse disorders, such as cancer, fibrosis, immune dysregulation, and neurodegenerative diseases. MMP-9, in particular, is highly dynamically regulated in several pathological processes. Development of MMP inhibitors has therefore been an attractive strategy for therapeutic intervention. However, a long history of failed clinical trials has demonstrated that broad-spectrum MMP inhibitors have limited clinical utility, which has spurred the development of inhibitors selective for individual MMPs. Attaining selectivity has been technically challenging because of sequence and structural conservation across the various MMPs. Here, through a biochemical and structural screening paradigm, we have identified JNJ0966, a highly selective compound that inhibited activation of MMP-9 zymogen and subsequent generation of catalytically active enzyme. JNJ0966 had no effect on MMP-1, MMP-2, MMP-3, MMP-9, or MMP-14 catalytic activity and did not inhibit activation of the highly related MMP-2 zymogen. The molecular basis for this activity was characterized as an interaction of JNJ0966 with a structural pocket in proximity to the MMP-9 zymogen cleavage site near Arg-106, which is distinct from the catalytic domain. JNJ0966 was efficacious in reducing disease severity in a mouse experimental autoimmune encephalomyelitis model, demonstrating the viability of this therapeutic approach. This discovery reveals an unprecedented pharmacological approach to MMP inhibition, providing an opportunity to improve selectivity of future clinical drug candidates. Targeting zymogen activation in this manner may also allow for pharmaceutical exploration of other enzymes previously viewed as intractable drug targets.
Keywords: allosteric regulation; drug action; drug discovery; enzyme inhibitor; enzyme processing; matrix metalloproteinase (MMP); pharmacology.
© 2017 by The American Society for Biochemistry and Molecular Biology, Inc.
Conflict of interest statement
During the time this work was performed, all authors were full-time employees of Janssen Research and Development
Figures
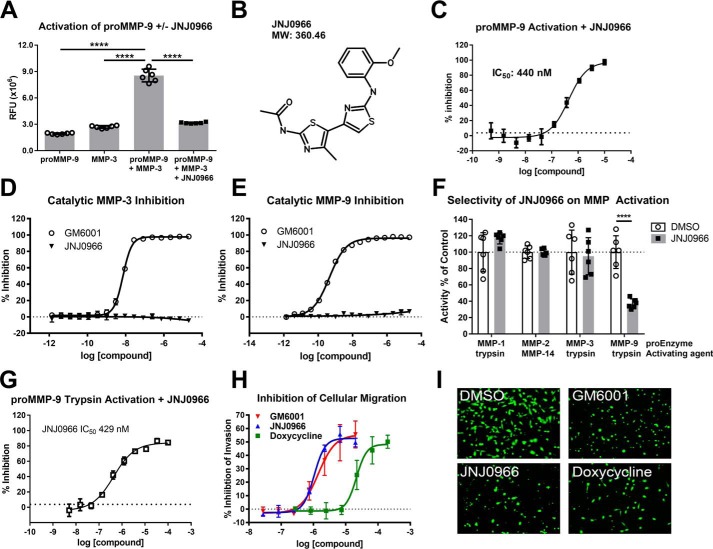
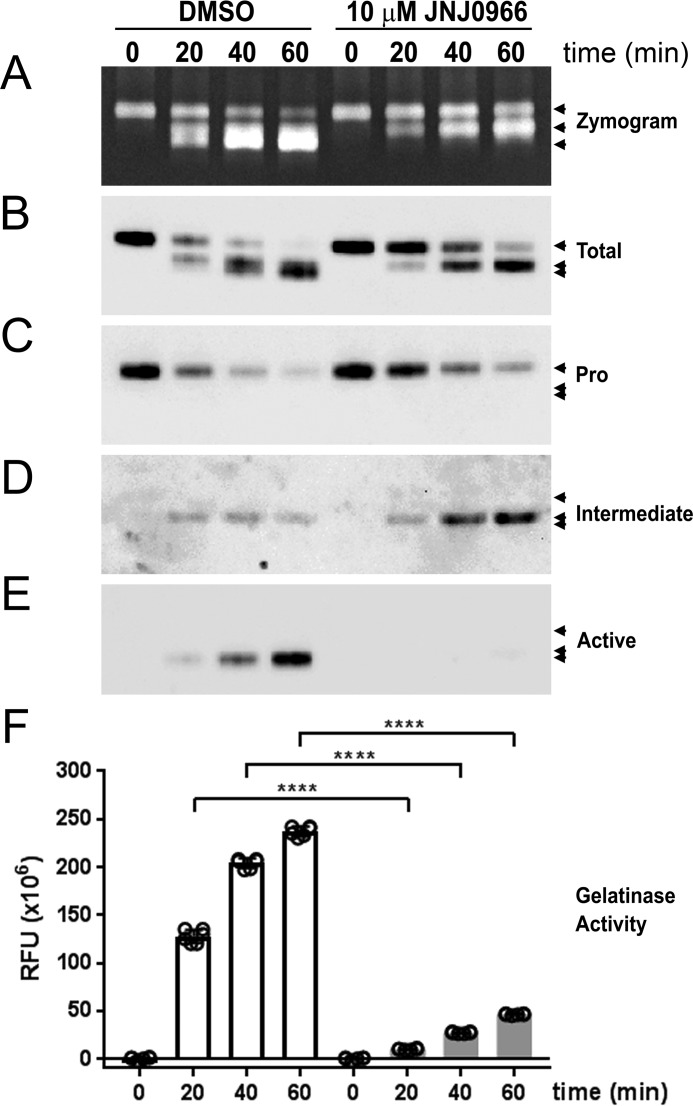
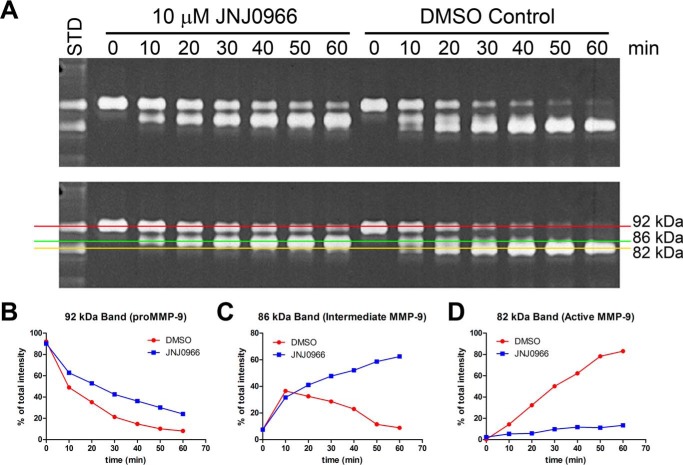
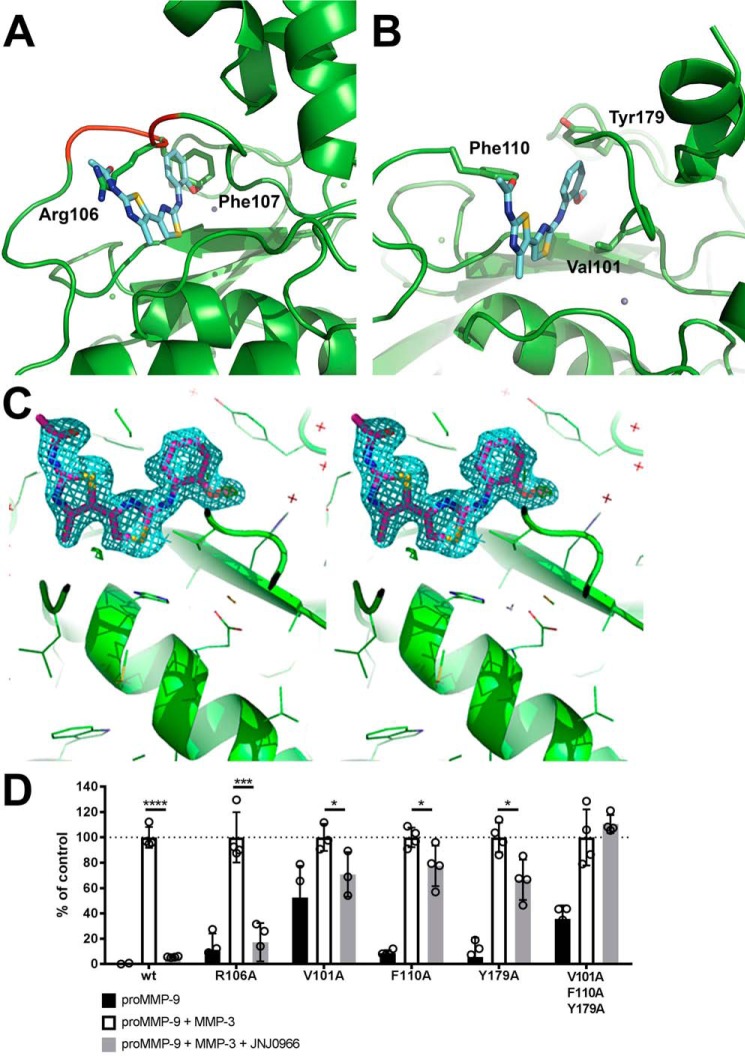
Comment in
-
Third time lucky? Getting a grip on matrix metalloproteinases.J Biol Chem. 2017 Oct 27;292(43):17975-17976. doi: 10.1074/jbc.H117.806075. J Biol Chem. 2017. PMID: 29079642 Free PMC article.
References
-
- Fingleton B. (2007) Matrix metalloproteinases as valid clinical targets. Curr. Pharm. Des. 13, 333–346 - PubMed
-
- Baranger K., Rivera S., Liechti F. D., Grandgirard D., Bigas J., Seco J., Tarrago T., Leib S. L., and Khrestchatisky M. (2014) Endogenous and synthetic MMP inhibitors in CNS physiopathology. Prog. Brain Res. 214, 313–351 - PubMed
-
- Golub L. M., Sorsa T., Lee H. M., Ciancio S., Sorbi D., Ramamurthy N. S., Gruber B., Salo T., and Konttinen Y. T. (1995) Doxycycline inhibits neutrophil (PMN)-type matrix metalloproteinases in human adult periodontitis gingiva. J. Clin. Periodontol. 22, 100–109 - PubMed
-
- Skiles J. W., Gonnella N. C., and Jeng A. Y. (2004) The design, structure, and clinical update of small molecular weight matrix metalloproteinase inhibitors. Curr. Med. Chem. 11, 2911–2977 - PubMed
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
