

Resetting of the Baroreflex Control of Sympathetic Vasomotor Activity during Natural Behaviors: Description and Conceptual Model of Central Mechanisms
- PMID: 28860965
- PMCID: PMC5559464
- DOI: 10.3389/fnins.2017.00461
Resetting of the Baroreflex Control of Sympathetic Vasomotor Activity during Natural Behaviors: Description and Conceptual Model of Central Mechanisms
Abstract
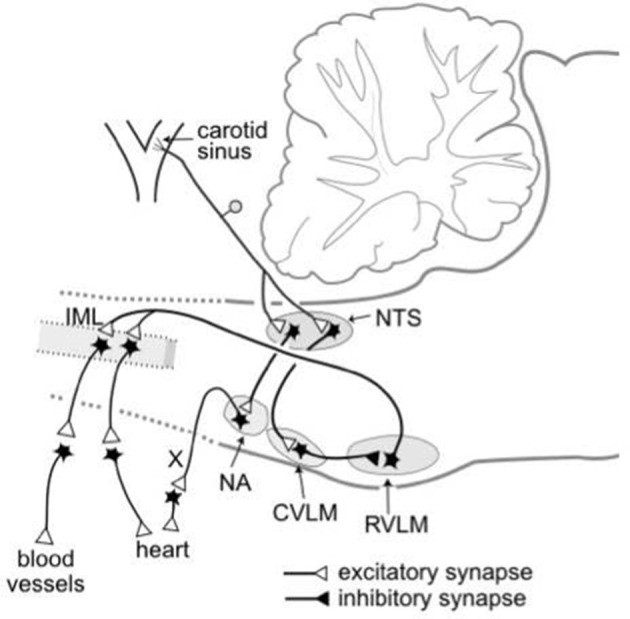
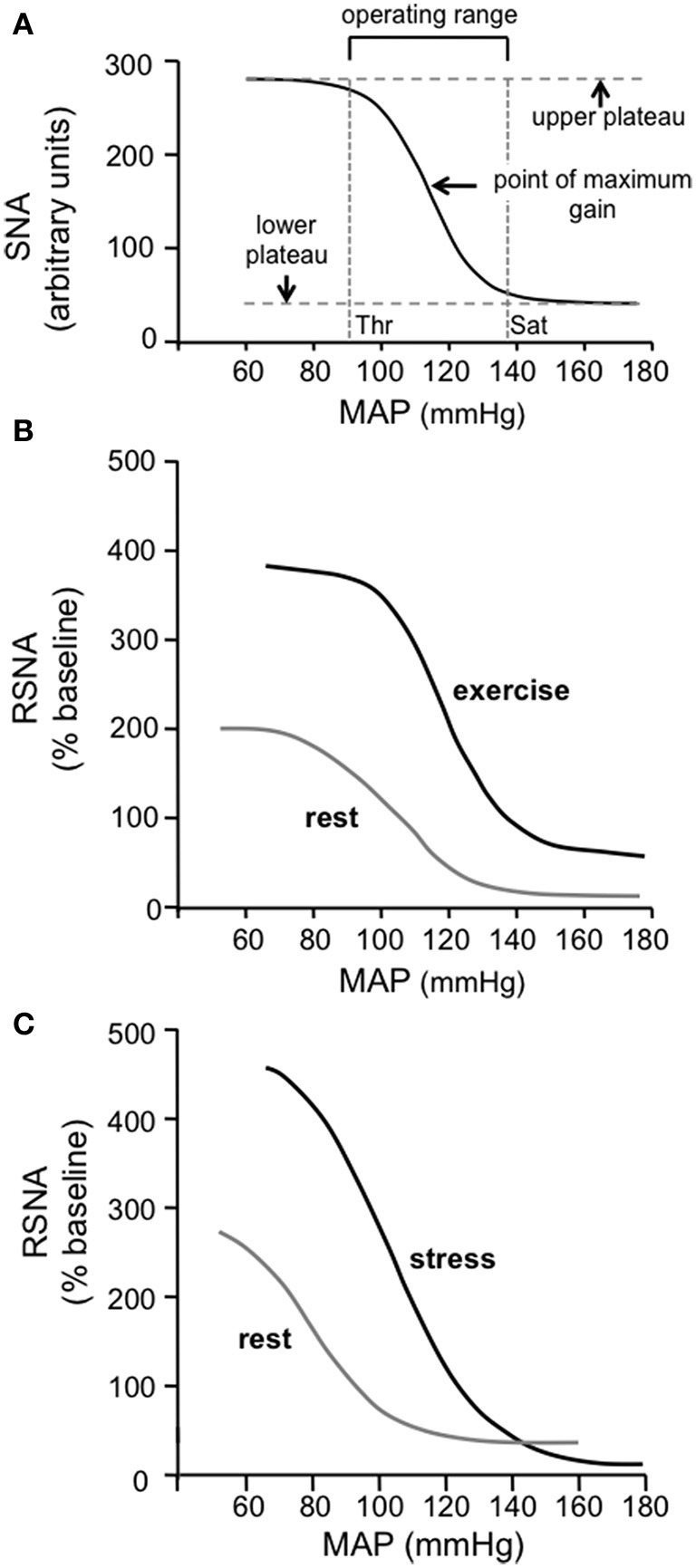
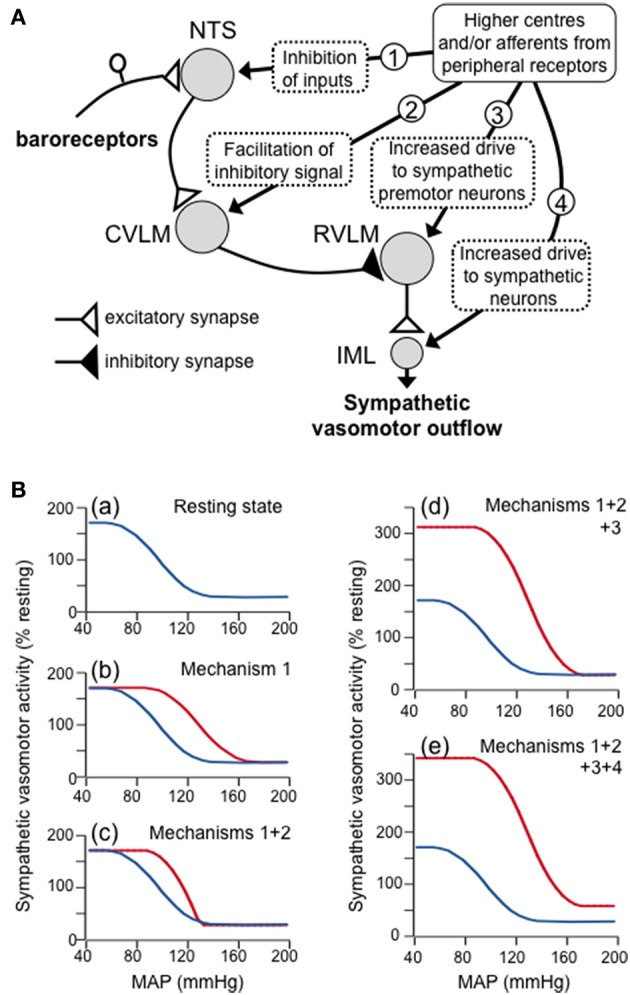
The baroreceptor reflex controls arterial pressure primarily via reflex changes in vascular resistance, rather than cardiac output. The vascular resistance in turn is dependent upon the activity of sympathetic vasomotor nerves innervating arterioles in different vascular beds. In this review, the major theme is that the baroreflex control of sympathetic vasomotor activity is not constant, but varies according to the behavioral state of the animal. In contrast to the view that was generally accepted up until the 1980s, I argue that the baroreflex control of sympathetic vasomotor activity is not inhibited or overridden during behaviors such as mental stress or exercise, but instead is reset under those conditions so that it continues to be highly effective in regulating sympathetic activity and arterial blood pressure at levels that are appropriate for the particular ongoing behavior. A major challenge is to identify the central mechanisms and neural pathways that subserve such resetting in different states. A model is proposed that is capable of simulating the different ways in which baroreflex resetting is occurred. Future studies are required to determine whether this proposed model is an accurate representation of the central mechanisms responsible for baroreflex resetting.
Keywords: baroreflex resetting; behavioral state; central baroreflex pathways; logistic function curves; models of baroreflex function; sympathetic vasomotor activity.
Figures
References
-
- Bailey T. W. (2006). Cranial visceral afferent pathways through the nucleus of the solitary tract to caudal ventrolateral medulla or paraventricular hypothalamus: target-specific synaptic reliability and convergence patterns. J. Neurosci. 26, 11893–11902. 10.1523/JNEUROSCI.2044-06.2006 - DOI - PMC - PubMed
-
- Burattini R., Socionovo G., Bellocchi F. (1994). On the approximation of static open-loop characteristics of baroreceptor reflex. Am. J. Physiol. 267, H267–H275. - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources