

Autoantibody Repertoire in APECED Patients Targets Two Distinct Subgroups of Proteins
- PMID: 28861084
- PMCID: PMC5561390
- DOI: 10.3389/fimmu.2017.00976
Autoantibody Repertoire in APECED Patients Targets Two Distinct Subgroups of Proteins
Abstract
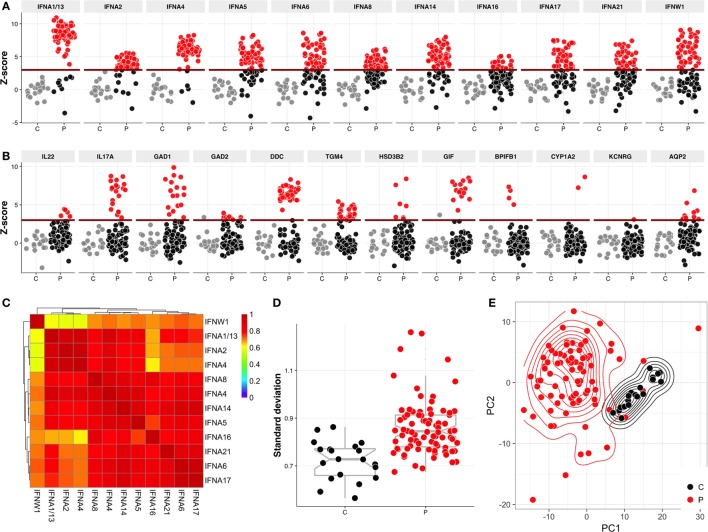
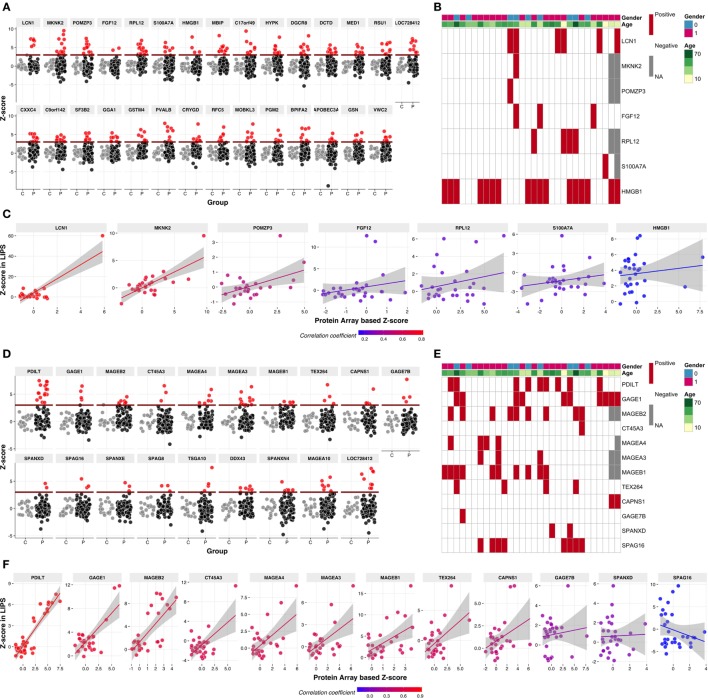
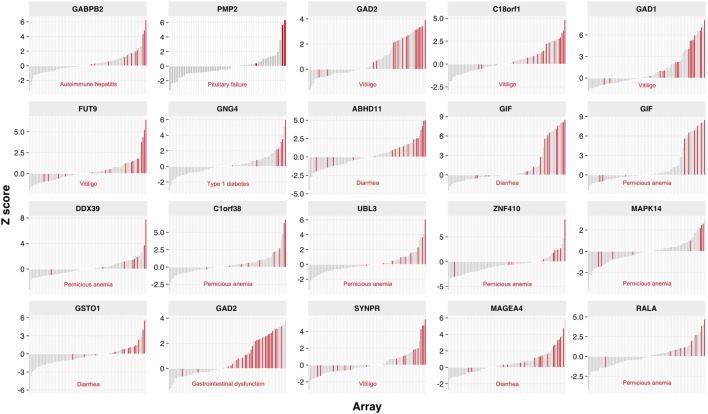
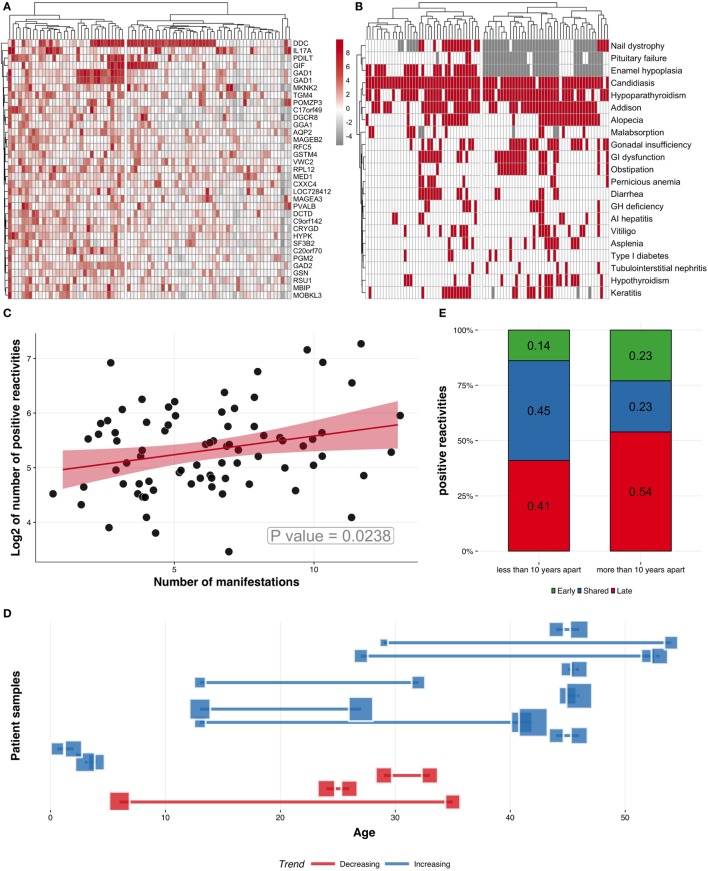
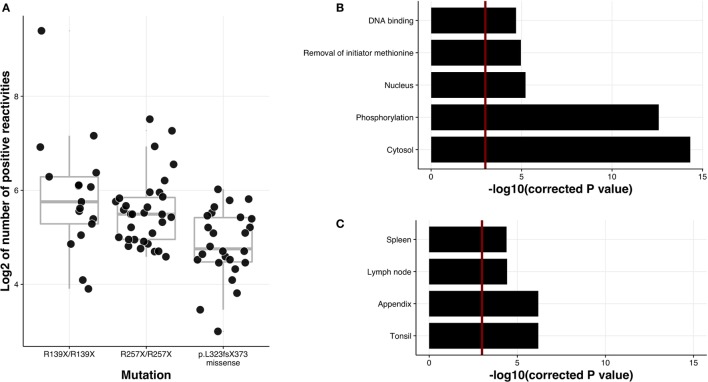
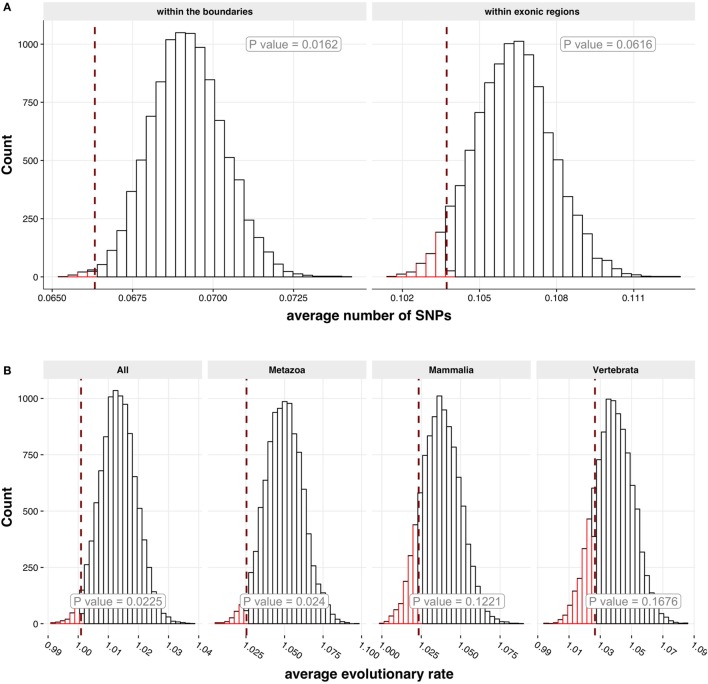
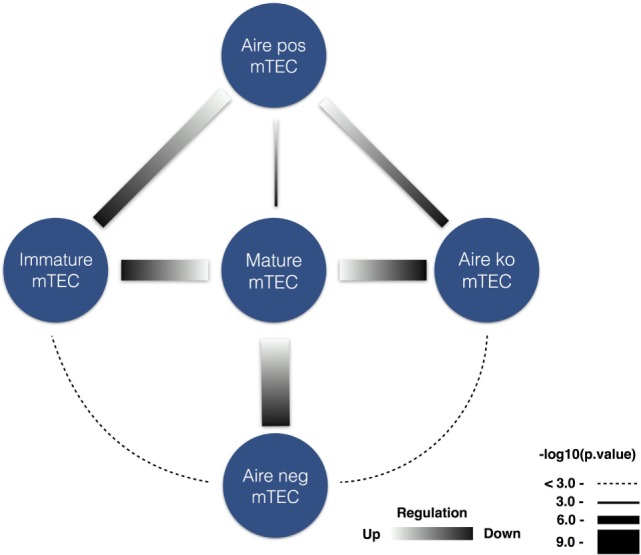
High titer autoantibodies produced by B lymphocytes are clinically important features of many common autoimmune diseases. APECED patients with deficient autoimmune regulator (AIRE) gene collectively display a broad repertoire of high titer autoantibodies, including some which are pathognomonic for major autoimmune diseases. AIRE deficiency severely reduces thymic expression of gene-products ordinarily restricted to discrete peripheral tissues, and developing T cells reactive to those gene-products are not inactivated during their development. However, the extent of the autoantibody repertoire in APECED and its relation to thymic expression of self-antigens are unclear. We here undertook a broad protein array approach to assess autoantibody repertoire in APECED patients. Our results show that in addition to shared autoantigen reactivities, APECED patients display high inter-individual variation in their autoantigen profiles, which collectively are enriched in evolutionarily conserved, cytosolic and nuclear phosphoproteins. The APECED autoantigens have two major origins; proteins expressed in thymic medullary epithelial cells and proteins expressed in lymphoid cells. These findings support the hypothesis that specific protein properties strongly contribute to the etiology of B cell autoimmunity.
Keywords: autoantibodies; autoantigen; autoimmune regulator; immune tolerance; thymus.
Figures
References
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources