

Mating in the Closest Living Relatives of Animals Is Induced by a Bacterial Chondroitinase
- PMID: 28867285
- PMCID: PMC5599222
- DOI: 10.1016/j.cell.2017.08.005
Mating in the Closest Living Relatives of Animals Is Induced by a Bacterial Chondroitinase
Abstract
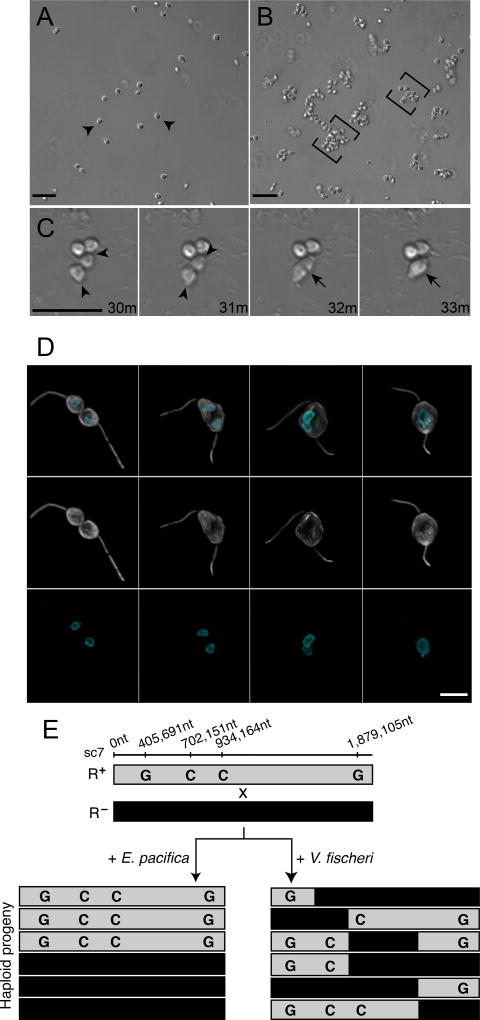
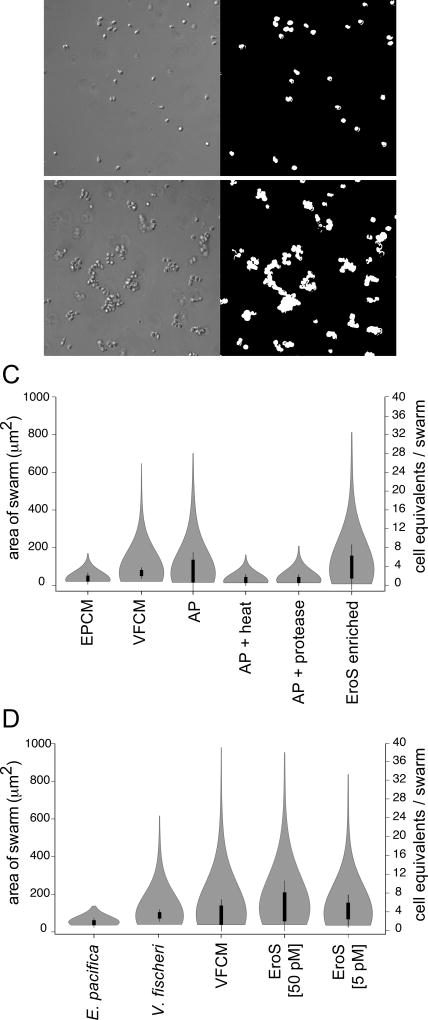
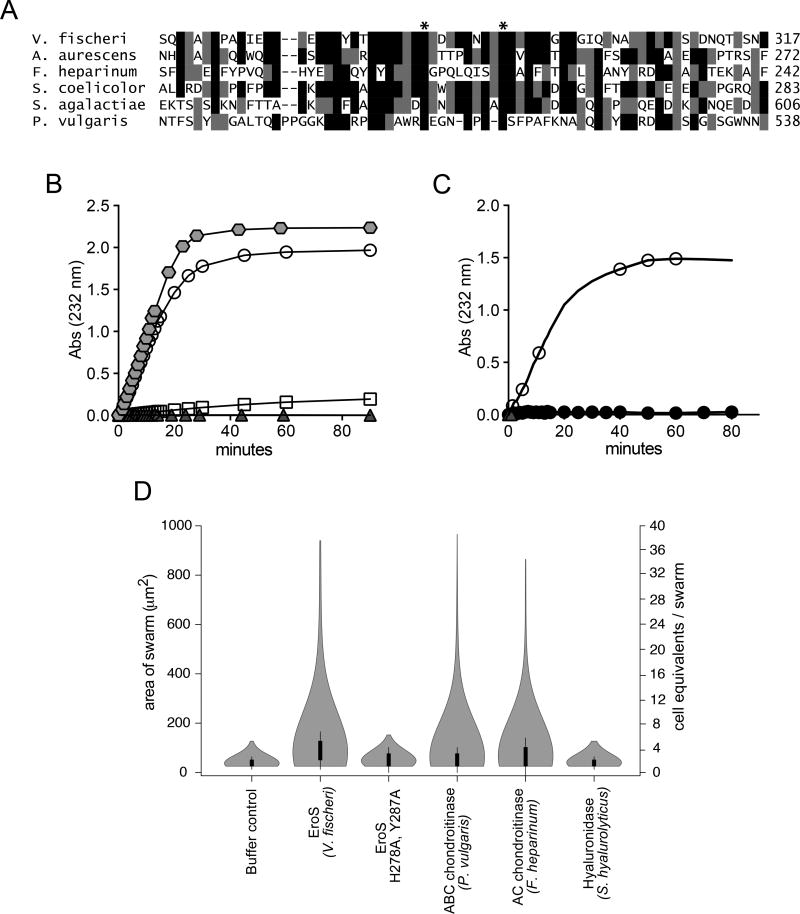
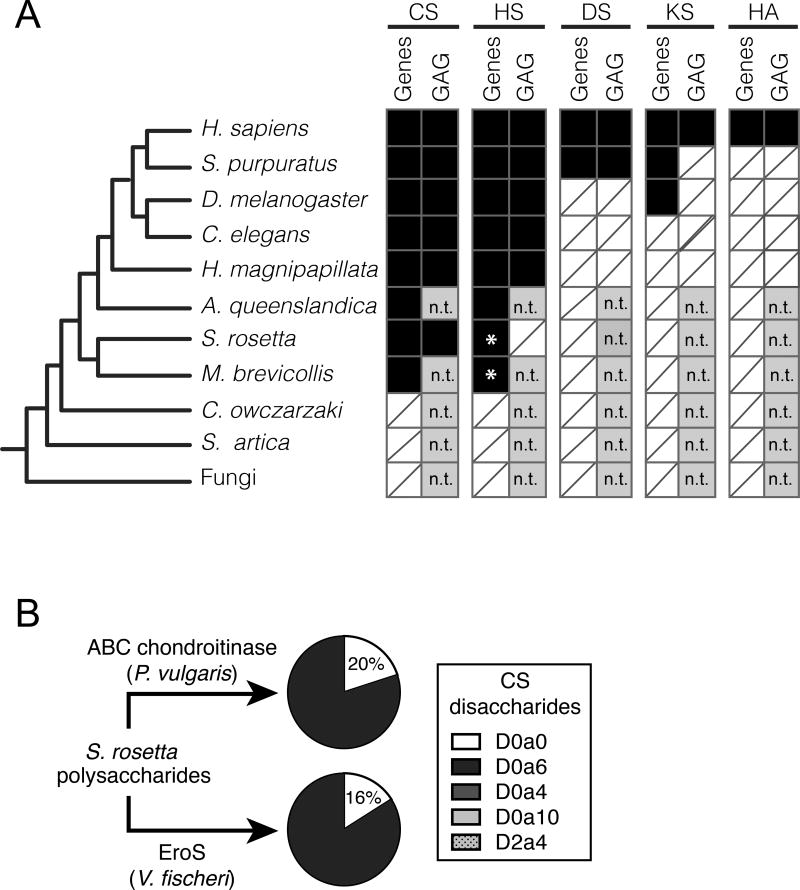
We serendipitously discovered that the marine bacterium Vibrio fischeri induces sexual reproduction in one of the closest living relatives of animals, the choanoflagellate Salpingoeca rosetta. Although bacteria influence everything from nutrition and metabolism to cell biology and development in eukaryotes, bacterial regulation of eukaryotic mating was unexpected. Here, we show that a single V. fischeri protein, the previously uncharacterized EroS, fully recapitulates the aphrodisiac-like activity of live V. fischeri. EroS is a chondroitin lyase; although its substrate, chondroitin sulfate, was previously thought to be an animal synapomorphy, we demonstrate that S. rosetta produces chondroitin sulfate and thus extend the ancestry of this important glycosaminoglycan to the premetazoan era. Finally, we show that V. fischeri, purified EroS, and other bacterial chondroitin lyases induce S. rosetta mating at environmentally relevant concentrations, suggesting that bacteria likely regulate choanoflagellate mating in nature.
Keywords: Vibrio; choanoflagellate; chondroitin lyase; chondroitin sulfate; chondroitinase; host-microbe; mating; swarming; symbiosis.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Eukaryotic Sexual Reproduction Evoked "with a Little Help from My Friends".Cell. 2017 Sep 7;170(6):1059-1061. doi: 10.1016/j.cell.2017.08.038. Cell. 2017. PMID: 28886378
References
-
- Ahn MY, Shin KH, Kim DH, Jung E-A, Toida T, Linhardt RJ, Kim YS. Characterization of a Bacteroides species from human intestine that degrades glycosaminoglycans. Can. J. Microbiol. 2011;44:423–429. - PubMed
-
- Avery MI. Lekking in birds: choice, competition and reproductive constraints. Ibis. 1984;126:177–187.
-
- Babcock R, Mundy C, Keesing J, Oliver J. Predictable and unpredictable spawning events: in situ behavioural data from free-spawning coral reef invertebrates. Invertebrate Reproduction & Development. 2011;22:213–227.
-
- Bartels-Hardege HD, Hardege JD, Zeeck E, Müller C, Wu BL, Zhu MY. Sex pheromones in marine polychaetes: a biologically active volatile compound from the coelomic fluid of female Nereis (Neanthes) japonica. Journal of Experimental Marine Biology and Ecology. 1996;201:275–284.
