

Evolutionary morphology of the lizard chemosensory system
- PMID: 28871144
- PMCID: PMC5583331
- DOI: 10.1038/s41598-017-09415-7
Evolutionary morphology of the lizard chemosensory system
Erratum in
-
Author Correction: Evolutionary morphology of the lizard chemosensory system.Sci Rep. 2017 Dec 11;7(1):17570. doi: 10.1038/s41598-017-17185-5. Sci Rep. 2017. PMID: 29229944 Free PMC article.
Abstract
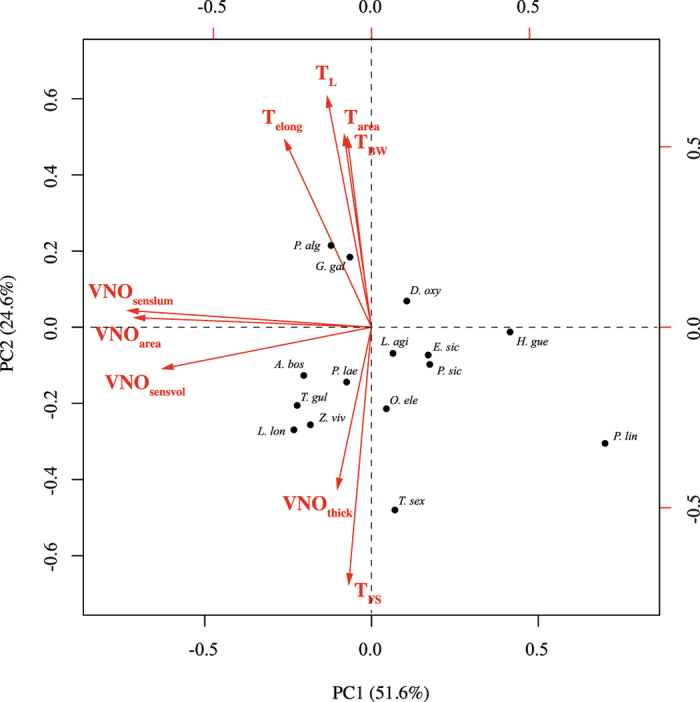
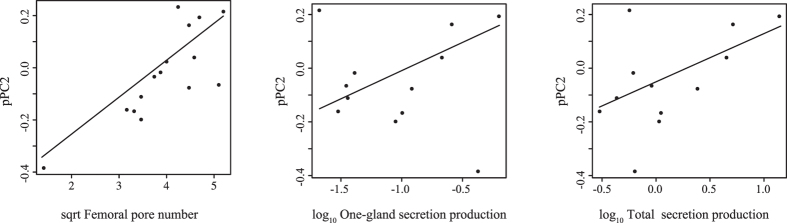
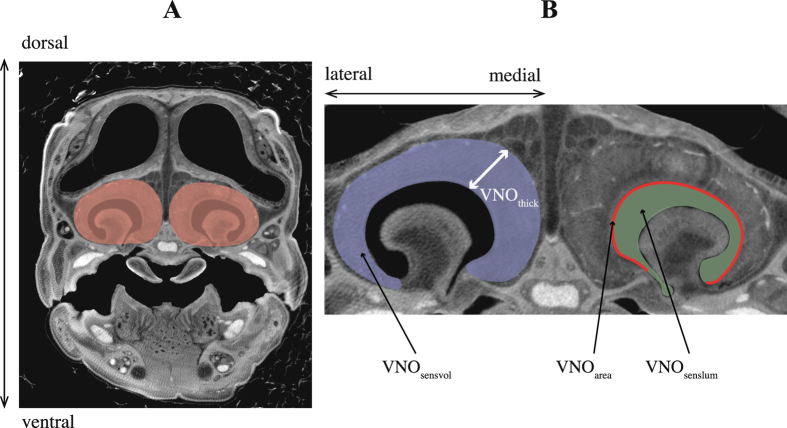
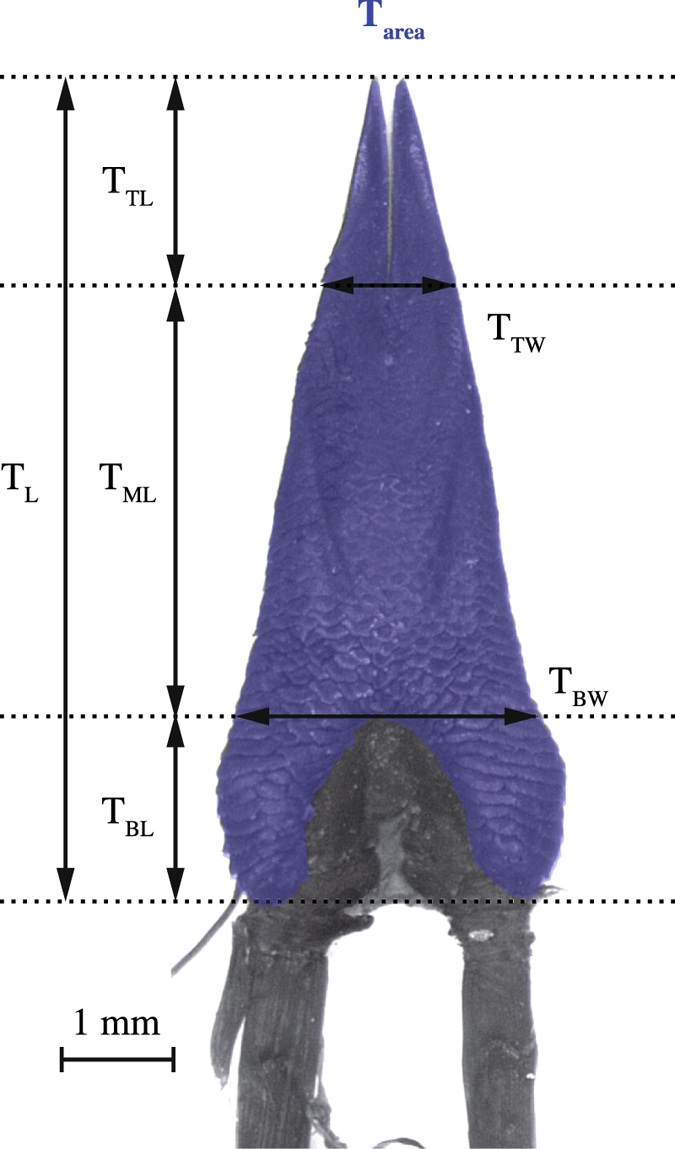
Foraging mode plays a pivotal role in traditional reconstructions of squamate evolution. Transitions between modes are said to spark concerted changes in the morphology, physiology, behaviour, and life history of lizards. With respect to their sensory systems, species that adopt a sit-and-wait strategy are thought to rely on visual cues primarily, while actively hunting species would predominantly use chemical information. The morphology of the tongue and the vomeronasal-organs is believed to mirror this dichotomy. Still, support for this idea of concerted evolution of the morphology of the lizard sensory system merely originates from studies comparing only a few, distantly related taxa that differ in many aspects of their biology besides foraging mode. Hence, we compared vomeronasal-lingual morphology among closely related lizard species (Lacertidae). Our findings show considerable interspecific variation indicating that the chemosensory system of lacertids has undergone substantial change over a short evolutionary time. Although our results imply independent evolution of tongue and vomeronasal-organ form, we find evidence for co-variation between sampler and sensor, hinting towards an 'optimization' for efficient chemoreception. Furthermore, our findings suggest species' degree of investment in chemical signalling, and not foraging behaviour, as a leading factor driving the diversity in vomeronasal-lingual morphology among lacertid species.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures


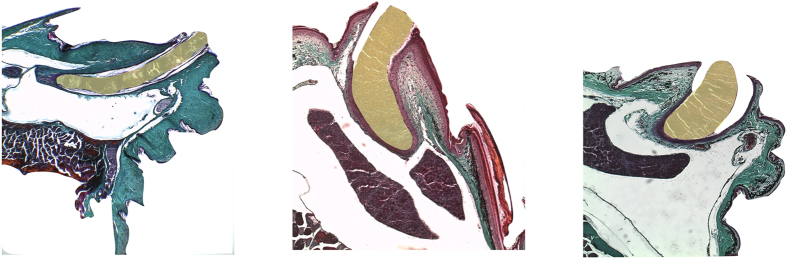
References
-
- Cooper WE. Variation and evolution of forked tongues in squamate reptiles. Herpetol. Nat. Hist. 1996;4:135–150.
-
- Cooper WE. Correlated evolution of prey chemical discrimination with foraging, lingual morphology and vomeronasal chemoreceptor abundance in lizards. Behav. Ecol. Sociobiol. 1997;41:257–265. doi: 10.1007/s002650050387. - DOI
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
