

Identification and Functional Characterization of Small Alarmone Synthetases in Corynebacterium glutamicum
- PMID: 28871248
- PMCID: PMC5566576
- DOI: 10.3389/fmicb.2017.01601
Identification and Functional Characterization of Small Alarmone Synthetases in Corynebacterium glutamicum
Abstract
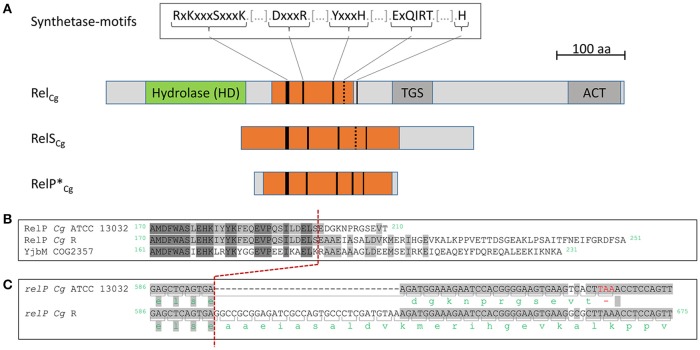
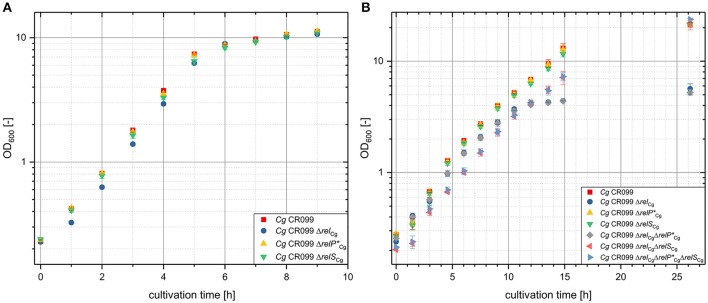
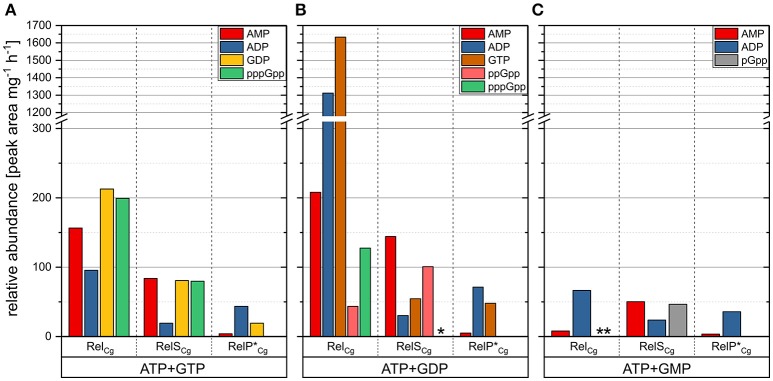
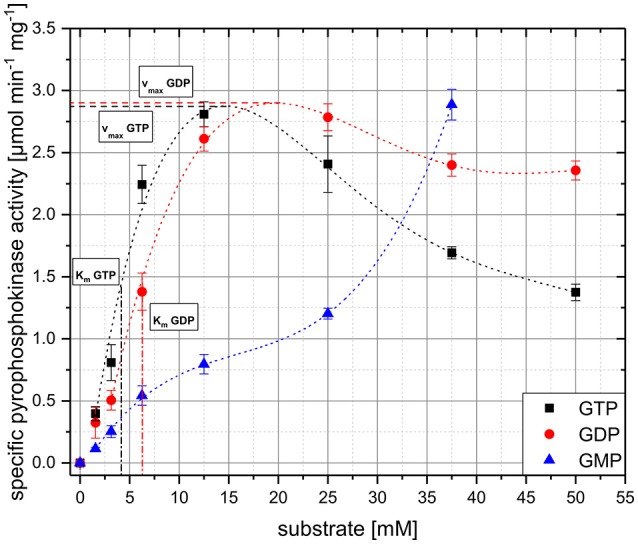
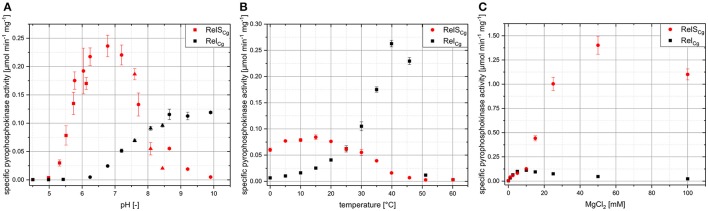
The hyperphosphorylated guanosine derivatives ppGpp and pppGpp represent global regulators of the bacterial stress response, as they act as central elements of the stringent response system. Although it was assumed that both, (p)ppGpp synthesis and hydrolysis, are catalyzed by one bifunctional RSH-protein in the actinobacterial model organism Corynebacterium glutamicum ATCC 13032, two putative short alarmone synthetases (SASs) were identified by bioinformatic analyses. The predicted sequences of both enzymes, designated as RelP*Cg and RelSCg, exhibit high similarities to the conserved (p)ppGpp synthetase catalytic domain. In the context of sequence analysis, significant differences were found between the RelP variants of different C. glutamicum isolates. In contrast to the bifunctional RelA/SpoT homolog (RSH) protein RelCg, whose gene deletion results in a reduced growth rate, no change in growth characteristics were observed for deletion mutants of the putative SAS proteins under standard growth conditions. The growth deficit of the Δrel strain could be restored by the additional deletion of the gene encoding RelSCg, which clearly indicates a functional relationship between both enzymes. The predicted pyrophosphokinase activity of RelSCg was demonstrated by means of genetic complementation of an Escherichia coli ΔrelAΔspoT strain. For the expression of RelP*Cg , as well as the slightly differing variant RelPCg from C. glutamicum AS1.542, no complementation was observed, concluding that both RelP versions possess no significant pyrophosphokinase activity in vivo. The results were confirmed by in vitro characterization of the corresponding proteins. In the course of this investigation, the additional conversion of GMP to pGpp was determined for the enzyme RelSCg. Since the SAS species analyzed extend both the network of stringent response related enzymes and the number of substances involved, the study of this class of enzymes is an important component in understanding the bacterial stress response. In addition to the comprehension of important biological processes, such as growth rate regulation and the survival of pathogenic species in the host organism, SAS enzymes can be used to produce novel hyperphosphorylated nucleotide species, such as pGpp.
Keywords: (p)ppGpp; ActRel; RelP; RelS; alarmone; pGpp; phylogeny; stringent response.
Figures

References
-
- Baumgart M., Unthan S., Ruckert C., Sivalingam J., Grunberger A., Kalinowski J., et al. (2013). Construction of a prophage-free variant of Corynebacterium glutamicum ATCC 13032 for use as a platform strain for basic research and industrial biotechnology. Appl. Environ. Microbiol. 79, 6006–6015. 10.1128/AEM.01634-13 - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases