

Use of a Yeast tRNase Killer Toxin to Diagnose Kti12 Motifs Required for tRNA Modification by Elongator
- PMID: 28872616
- PMCID: PMC5618205
- DOI: 10.3390/toxins9090272
Use of a Yeast tRNase Killer Toxin to Diagnose Kti12 Motifs Required for tRNA Modification by Elongator
Abstract
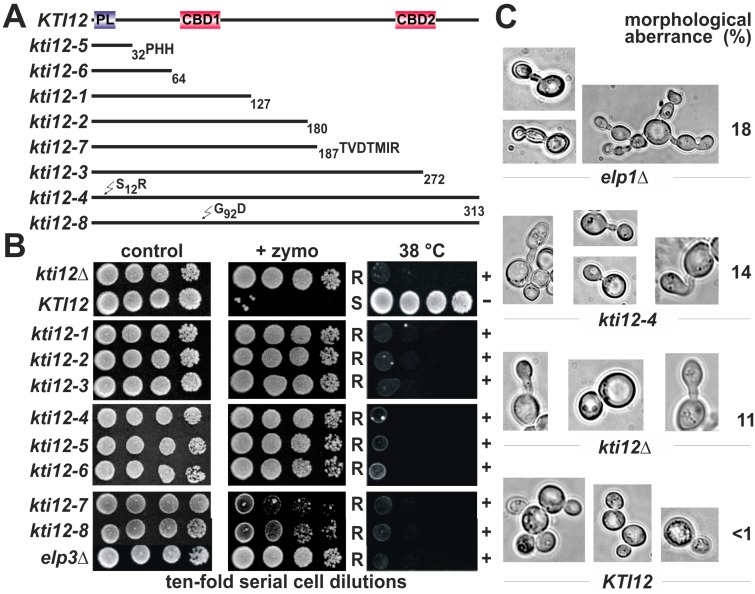
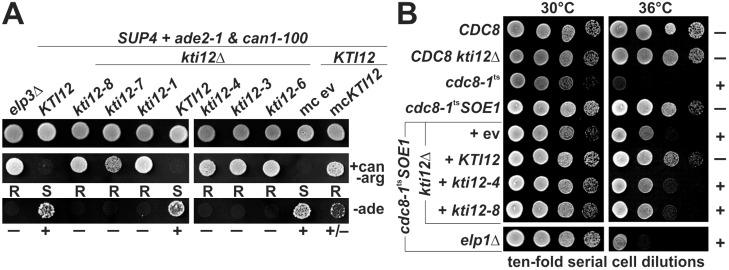
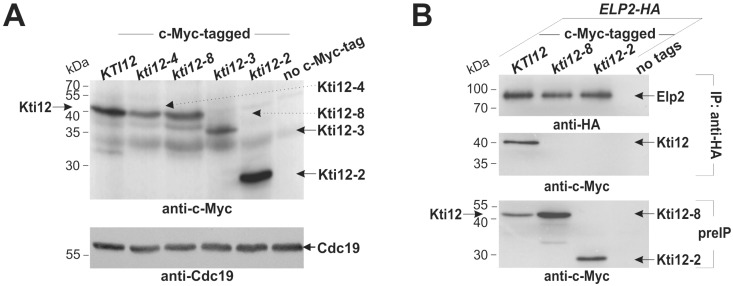
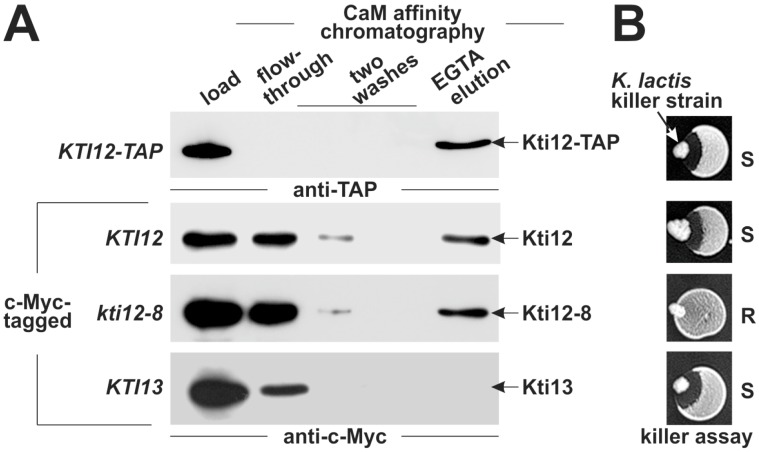
Saccharomyces cerevisiae cells are killed by zymocin, a tRNase ribotoxin complex from Kluyveromyces lactis, which cleaves anticodons and inhibits protein synthesis. Zymocin's action requires specific chemical modification of uridine bases in the anticodon wobble position (U34) by the Elongator complex (Elp1-Elp6). Hence, loss of anticodon modification in mutants lacking Elongator or related KTI (K. lactis Toxin Insensitive) genes protects against tRNA cleavage and confers resistance to the toxin. Here, we show that zymocin can be used as a tool to genetically analyse KTI12, a gene previously shown to code for an Elongator partner protein. From a kti12 mutant pool of zymocin survivors, we identify motifs in Kti12 that are functionally directly coupled to Elongator activity. In addition, shared requirement of U34 modifications for nonsense and missense tRNA suppression (SUP4; SOE1) strongly suggests that Kti12 and Elongator cooperate to assure proper tRNA functioning. We show that the Kti12 motifs are conserved in plant ortholog DRL1/ELO4 from Arabidopsis thaliana and seem to be involved in binding of cofactors (e.g., nucleotides, calmodulin). Elongator interaction defects triggered by mutations in these motifs correlate with phenotypes typical for loss of U34 modification. Thus, tRNA modification by Elongator appears to require physical contact with Kti12, and our preliminary data suggest that metabolic signals may affect proper communication between them.
Keywords: Elongator complex; Kti12; ribotoxin; tRNA anticodon modification; tRNase; zymocin.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
tRNA binding to Kti12 is crucial for wobble uridine modification by Elongator.Nucleic Acids Res. 2025 Apr 10;53(7):gkaf296. doi: 10.1093/nar/gkaf296. Nucleic Acids Res. 2025. PMID: 40226916 Free PMC article.
-
tRNA and protein methylase complexes mediate zymocin toxicity in yeast.Mol Microbiol. 2008 Sep;69(5):1266-77. doi: 10.1111/j.1365-2958.2008.06358.x. Epub 2008 Jul 24. Mol Microbiol. 2008. PMID: 18657261
-
Elongator function depends on antagonistic regulation by casein kinase Hrr25 and protein phosphatase Sit4.Mol Microbiol. 2009 Sep;73(5):869-81. doi: 10.1111/j.1365-2958.2009.06811.x. Epub 2009 Jul 28. Mol Microbiol. 2009. PMID: 19656297
-
Zymocin, a composite chitinase and tRNase killer toxin from yeast.Biochem Soc Trans. 2007 Dec;35(Pt 6):1533-7. doi: 10.1042/BST0351533. Biochem Soc Trans. 2007. PMID: 18031261 Review.
-
Elongator, a conserved complex required for wobble uridine modifications in eukaryotes.RNA Biol. 2014;11(12):1519-28. doi: 10.4161/15476286.2014.992276. RNA Biol. 2014. PMID: 25607684 Free PMC article. Review.
Cited by
-
Kti12, a PSTK-like tRNA dependent ATPase essential for tRNA modification by Elongator.Nucleic Acids Res. 2019 May 21;47(9):4814-4830. doi: 10.1093/nar/gkz190. Nucleic Acids Res. 2019. PMID: 30916349 Free PMC article.
-
Comparative Analysis of Proteins Regulated during Cadmium Sulfide Quantum Dots Response in Arabidopsis thaliana Wild Type and Tolerant Mutants.Nanomaterials (Basel). 2021 Mar 1;11(3):615. doi: 10.3390/nano11030615. Nanomaterials (Basel). 2021. PMID: 33804515 Free PMC article.
-
Collaboration of tRNA modifications and elongation factor eEF1A in decoding and nonsense suppression.Sci Rep. 2018 Aug 24;8(1):12749. doi: 10.1038/s41598-018-31158-2. Sci Rep. 2018. PMID: 30143741 Free PMC article.
-
Evolutionary Conservation in Protein-Protein Interactions and Structures of the Elongator Sub-Complex ELP456 from Arabidopsis and Yeast.Int J Mol Sci. 2024 Apr 15;25(8):4370. doi: 10.3390/ijms25084370. Int J Mol Sci. 2024. PMID: 38673955 Free PMC article.
-
Overexpression of GhKTI12 Enhances Seed Yield and Biomass Production in Nicotiana Tabacum.Genes (Basel). 2022 Feb 25;13(3):426. doi: 10.3390/genes13030426. Genes (Basel). 2022. PMID: 35327981 Free PMC article.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
