

Treg-specific IL-27Rα deletion uncovers a key role for IL-27 in Treg function to control autoimmunity
- PMID: 28874534
- PMCID: PMC5617261
- DOI: 10.1073/pnas.1703100114
Treg-specific IL-27Rα deletion uncovers a key role for IL-27 in Treg function to control autoimmunity
Abstract
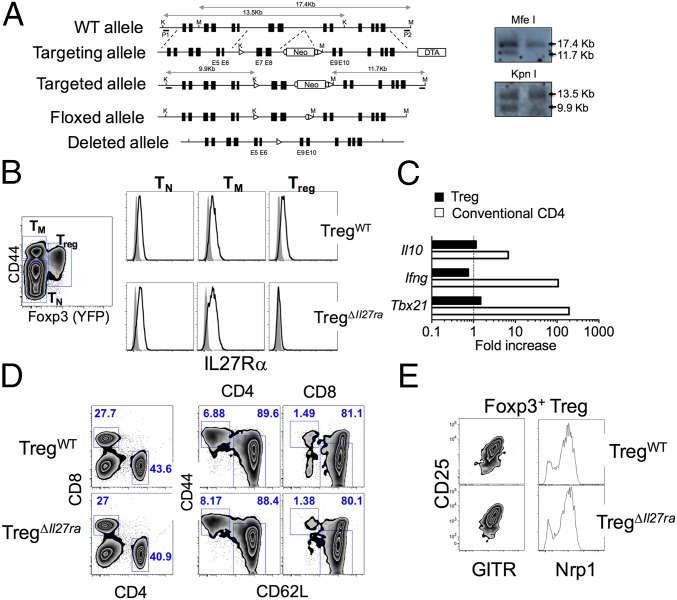
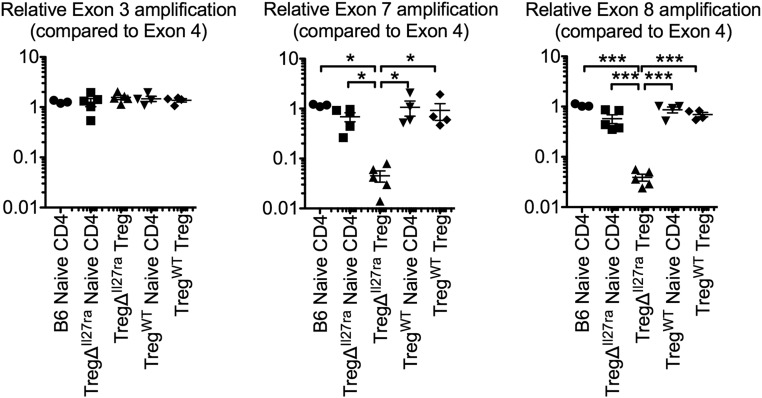
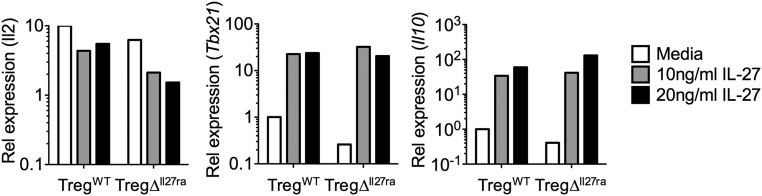
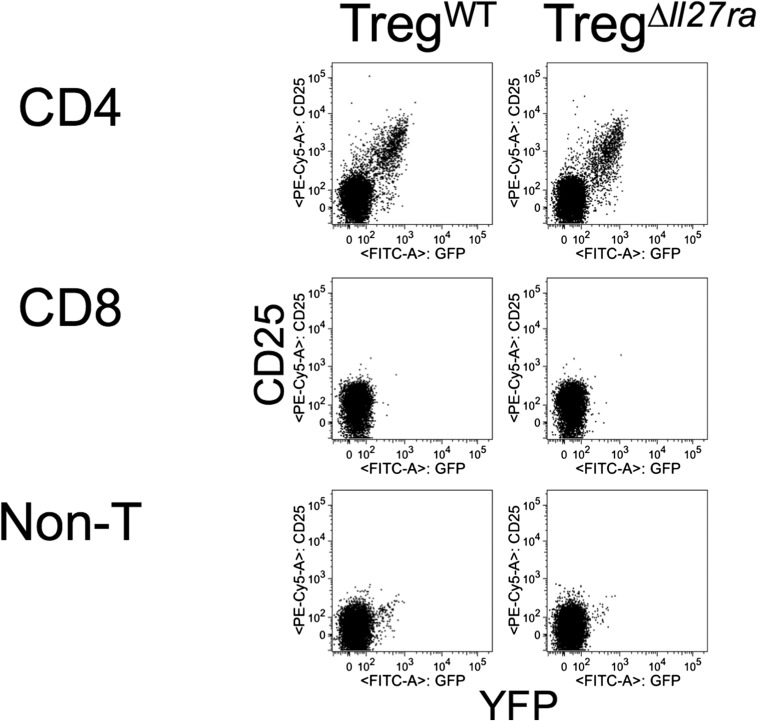
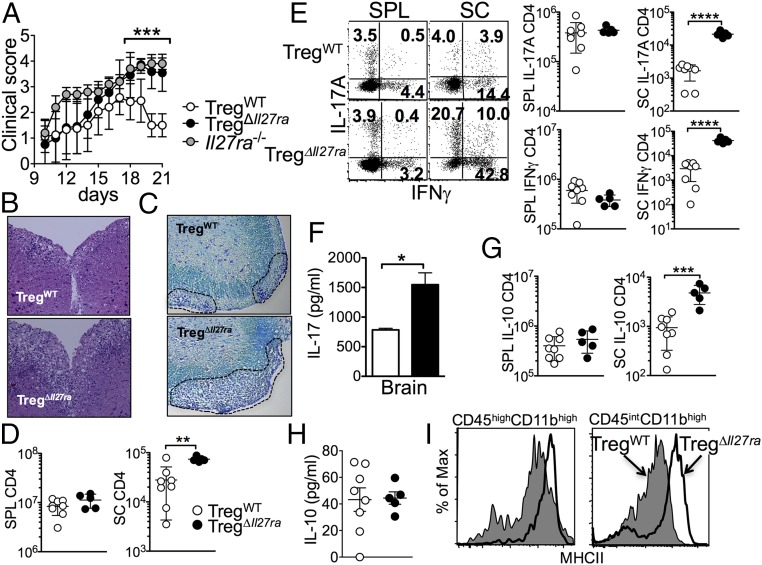
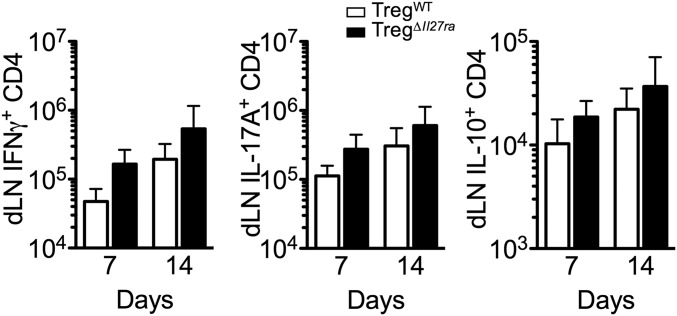
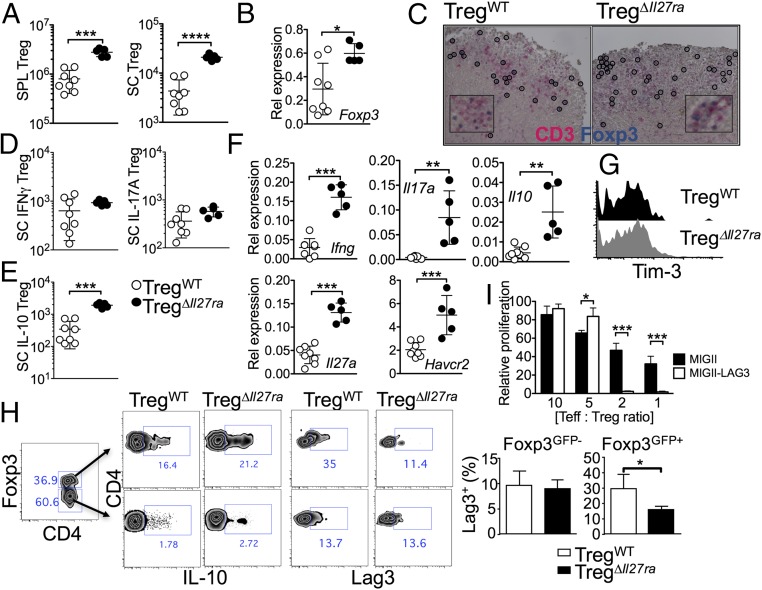
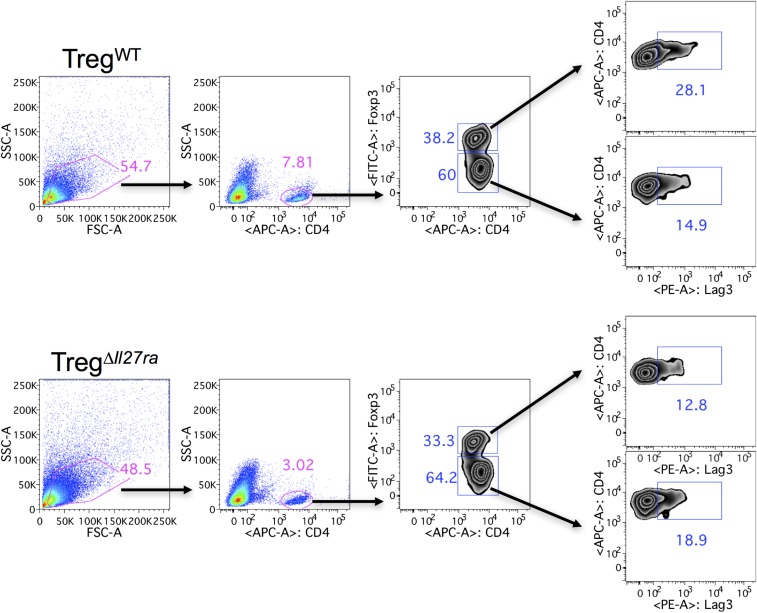
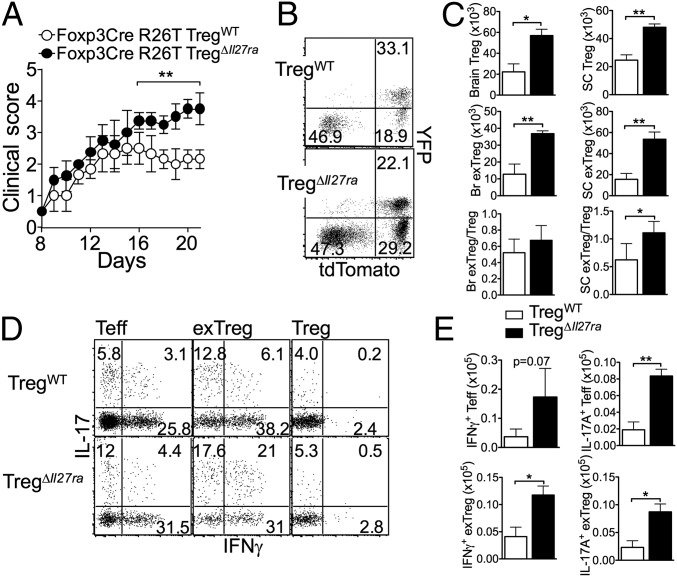
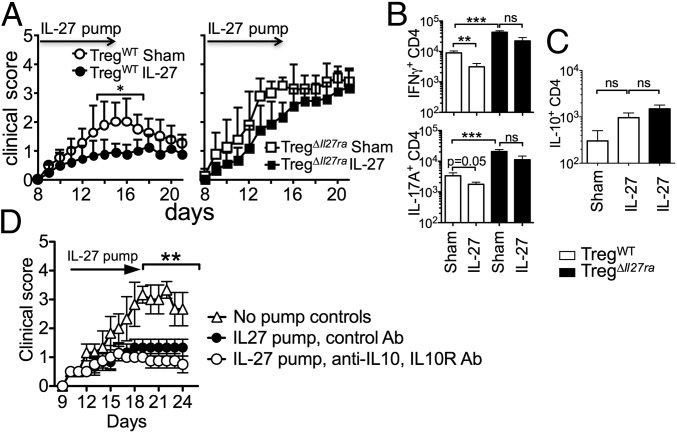
Dysregulated Foxp3+ Treg functions result in uncontrolled immune activation and autoimmunity. Therefore, identifying cellular factors modulating Treg functions is an area of great importance. Here, using Treg-specific Il27ra-/- mice, we report that IL-27 signaling in Foxp3+ Tregs is essential for Tregs to control autoimmune inflammation in the central nervous system (CNS). Following experimental autoimmune encephalomyelitis (EAE) induction, Treg-specific Il27ra-/- mice develop more severe EAE. Consistent with the severe disease, the numbers of IFNγ- and IL-17-producing CD4 T cells infiltrating the CNS tissues are greater in these mice. Treg accumulation in the inflamed CNS tissues is not affected by the lack of IL-27 signaling in Tregs, suggesting a functional defect of Il27ra-/- Tregs. IL-10 production by conventional CD4 T cells and their CNS accumulation are rather elevated in Treg-specific Il27ra-/- mice. Analysis with Treg fate-mapping reporter mice further demonstrates that IL-27 signaling in Tregs may control stability of Foxp3 expression. Finally, systemic administration of recombinant IL-27 in Treg-specific Il27ra-/- mice fails to ameliorate the disease even in the presence of IL-27-responsive conventional CD4 T cells. These findings uncover a previously unknown role of IL-27 in regulating Treg function to control autoimmune inflammation.
Keywords: Foxp3+ regulatory T cells; IL-27; Tr1 cells; autoimmunity.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
