

Improved base excision repair inhibition and bacteriophage Mu Gam protein yields C:G-to-T:A base editors with higher efficiency and product purity
- PMID: 28875174
- PMCID: PMC5576876
- DOI: 10.1126/sciadv.aao4774
Improved base excision repair inhibition and bacteriophage Mu Gam protein yields C:G-to-T:A base editors with higher efficiency and product purity
Abstract
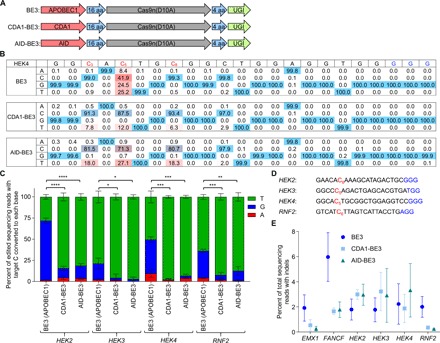
We recently developed base editing, the programmable conversion of target C:G base pairs to T:A without inducing double-stranded DNA breaks (DSBs) or requiring homology-directed repair using engineered fusions of Cas9 variants and cytidine deaminases. Over the past year, the third-generation base editor (BE3) and related technologies have been successfully used by many researchers in a wide range of organisms. The product distribution of base editing-the frequency with which the target C:G is converted to mixtures of undesired by-products, along with the desired T:A product-varies in a target site-dependent manner. We characterize determinants of base editing outcomes in human cells and establish that the formation of undesired products is dependent on uracil N-glycosylase (UNG) and is more likely to occur at target sites containing only a single C within the base editing activity window. We engineered CDA1-BE3 and AID-BE3, which use cytidine deaminase homologs that increase base editing efficiency for some sequences. On the basis of these observations, we engineered fourth-generation base editors (BE4 and SaBE4) that increase the efficiency of C:G to T:A base editing by approximately 50%, while halving the frequency of undesired by-products compared to BE3. Fusing BE3, BE4, SaBE3, or SaBE4 to Gam, a bacteriophage Mu protein that binds DSBs greatly reduces indel formation during base editing, in most cases to below 1.5%, and further improves product purity. BE4, SaBE4, BE4-Gam, and SaBE4-Gam represent the state of the art in C:G-to-T:A base editing, and we recommend their use in future efforts.
Figures





References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
