

Co-expression of xenopsin and rhabdomeric opsin in photoreceptors bearing microvilli and cilia
- PMID: 28876222
- PMCID: PMC5648526
- DOI: 10.7554/eLife.23435
Co-expression of xenopsin and rhabdomeric opsin in photoreceptors bearing microvilli and cilia
Abstract
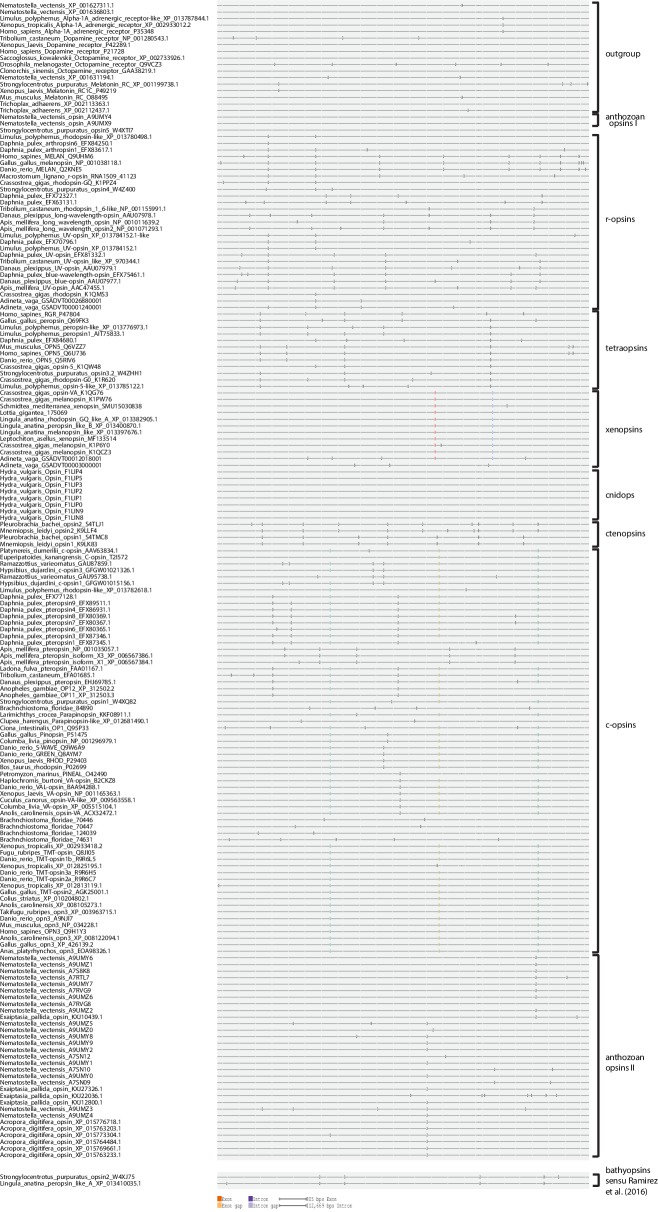
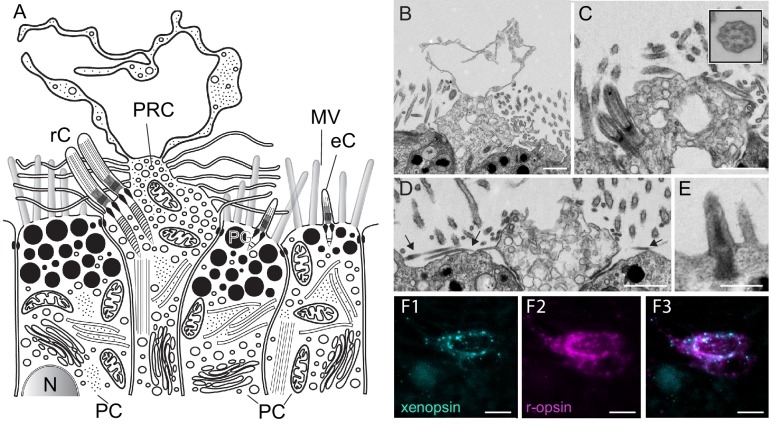
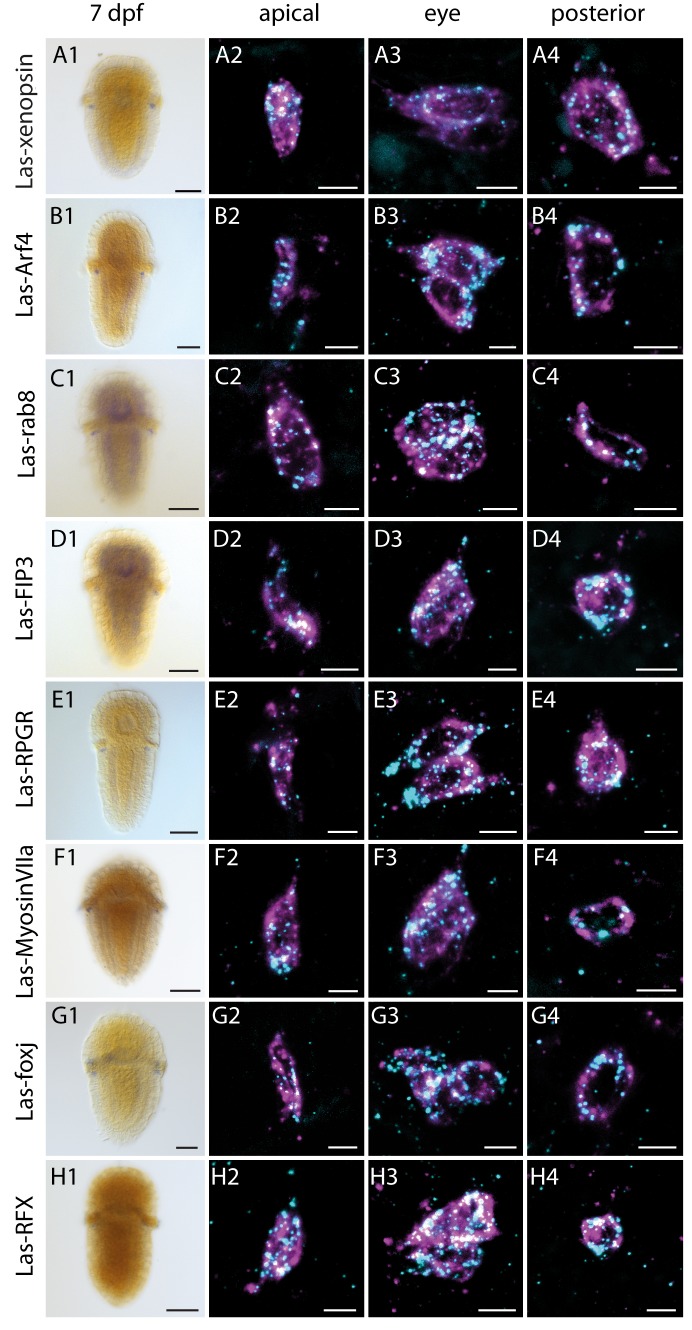
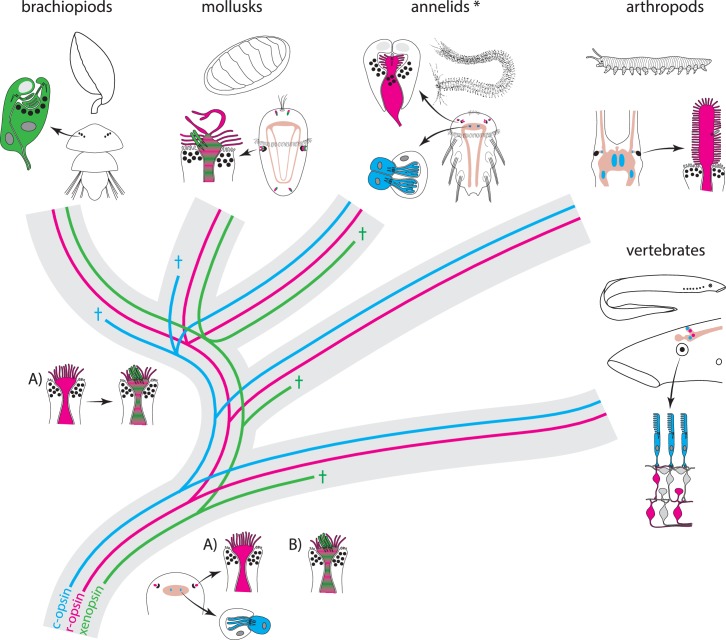
Ciliary and rhabdomeric opsins are employed by different kinds of photoreceptor cells, such as ciliary vertebrate rods and cones or protostome microvillar eye photoreceptors, that have specialized structures and molecular physiologies. We report unprecedented cellular co-expression of rhabdomeric opsin and a visual pigment of the recently described xenopsins in larval eyes of a mollusk. The photoreceptors bear both microvilli and cilia and express proteins that are orthologous to transporters in microvillar and ciliary opsin trafficking. Highly conserved but distinct gene structures suggest that xenopsins and ciliary opsins are of independent origin, irrespective of their mutually exclusive distribution in animals. Furthermore, we propose that frequent opsin gene loss had a large influence on the evolution, organization and function of brain and eye photoreceptor cells in bilaterian animals. The presence of xenopsin in eyes of even different design might be due to a common origin and initial employment of this protein in a highly plastic photoreceptor cell type of mixed microvillar/ciliary organization.
Keywords: Leptochiton asellus; evolution; evolutionary biology; eye; genomics; invertebrates; neuroscience; opsin; photoreceptor.
Conflict of interest statement
No competing interests declared.
Figures
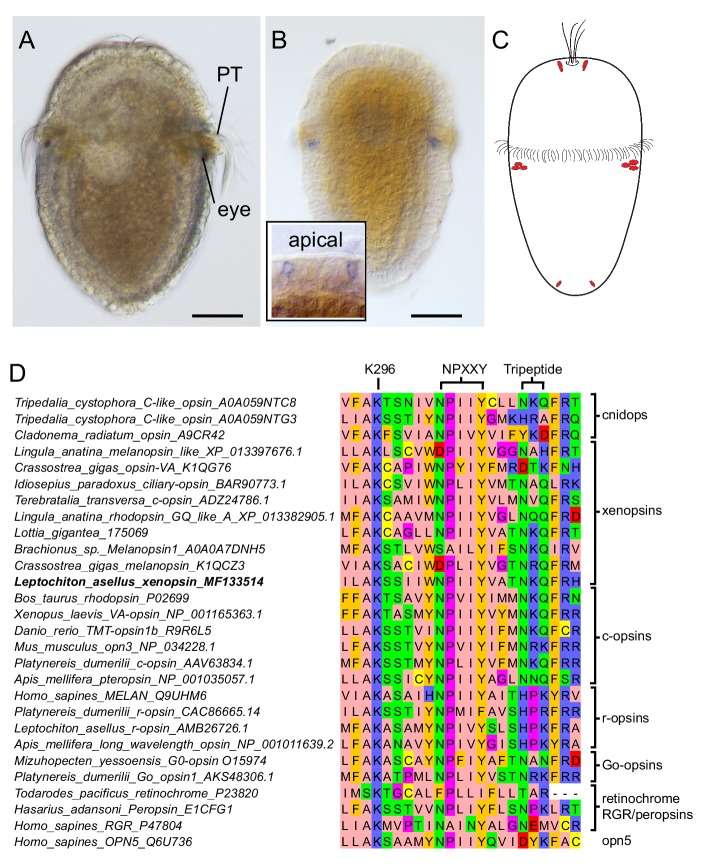

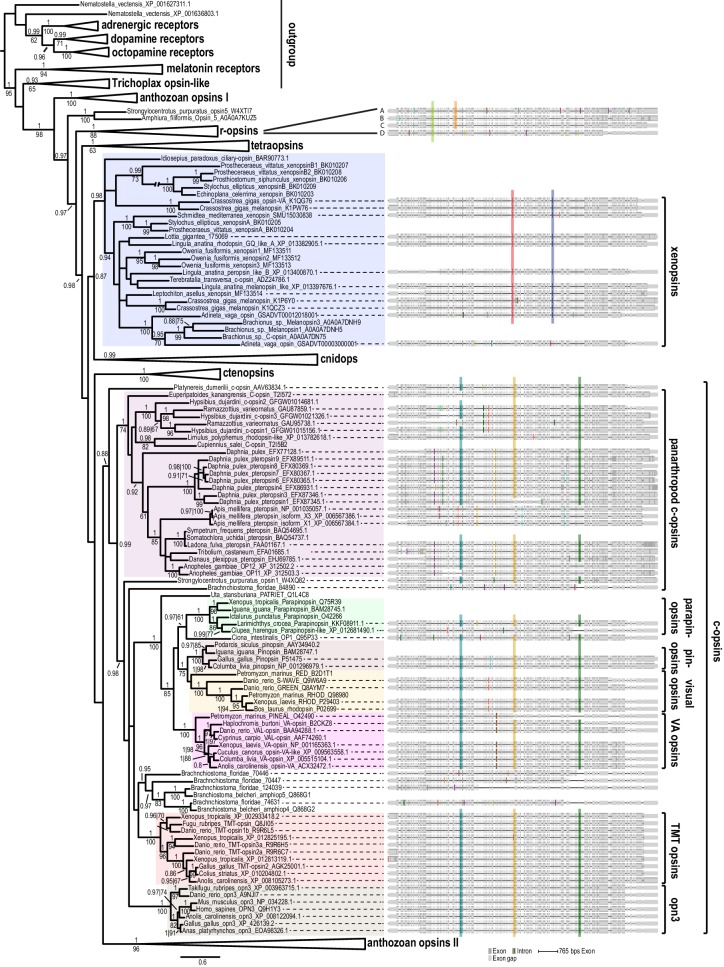
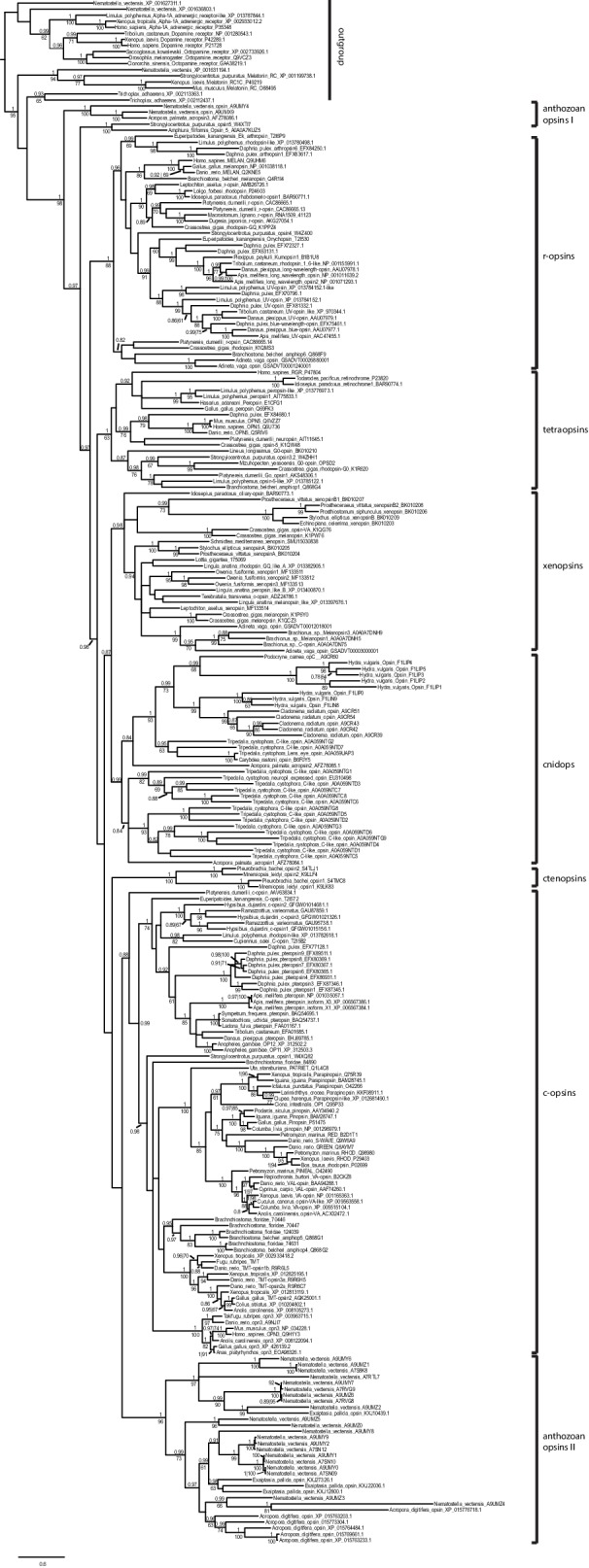
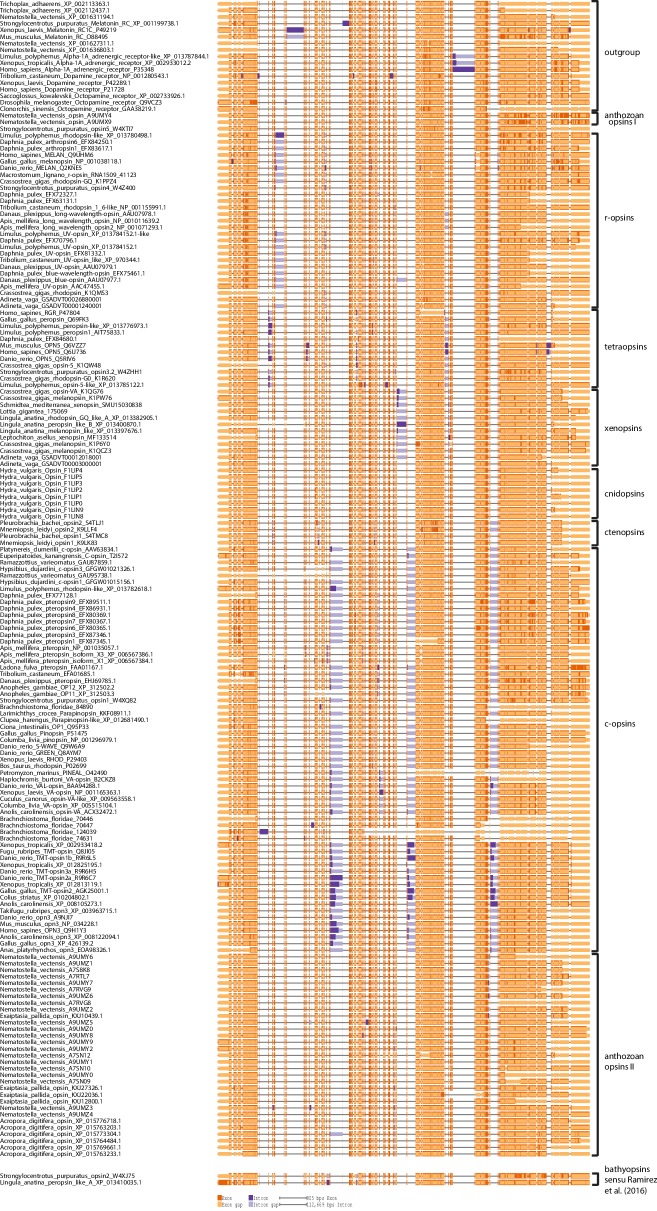
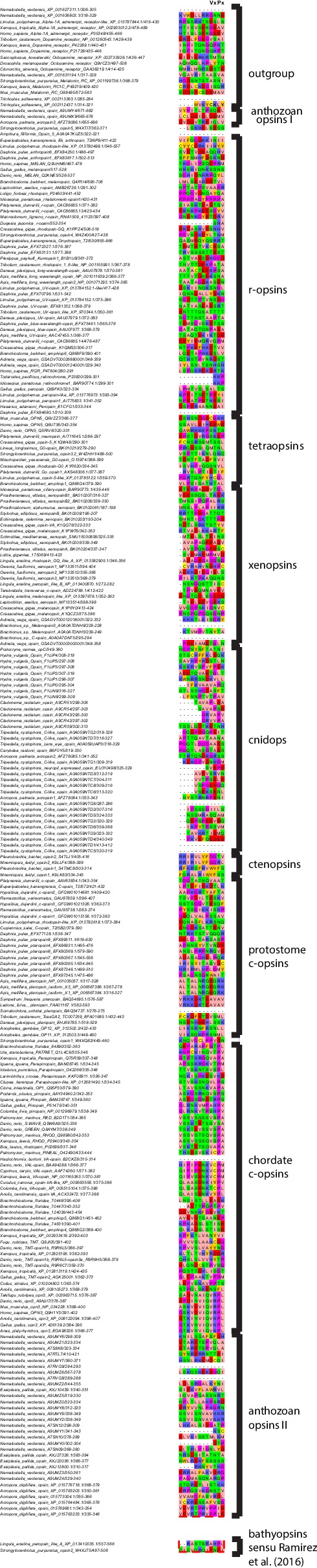
Comment in
-
The enigmatic xenopsins.Elife. 2017 Oct 19;6:e31781. doi: 10.7554/eLife.31781. Elife. 2017. PMID: 29048325 Free PMC article.
References
-
- Applebury ML, Antoch MP, Baxter LC, Chun LL, Falk JD, Farhangfar F, Kage K, Krzystolik MG, Lyass LA, Robbins JT. The murine cone photoreceptor: a single cone type expresses both S and M opsins with retinal spatial patterning. Neuron. 2000;27:513–523. - PubMed
-
- Arendt D, Tessmar K, de Campos-Baptista MI, Dorresteijn A, Wittbrodt J. Development of pigment-cup eyes in the polychaete Platynereis dumerilii and evolutionary conservation of larval eyes in Bilateria. Development. 2002a;129:1143–1154. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
