

New class of precision antimicrobials redefines role of Clostridium difficile S-layer in virulence and viability
- PMID: 28878013
- PMCID: PMC5603275
- DOI: 10.1126/scitranslmed.aah6813
New class of precision antimicrobials redefines role of Clostridium difficile S-layer in virulence and viability
Abstract
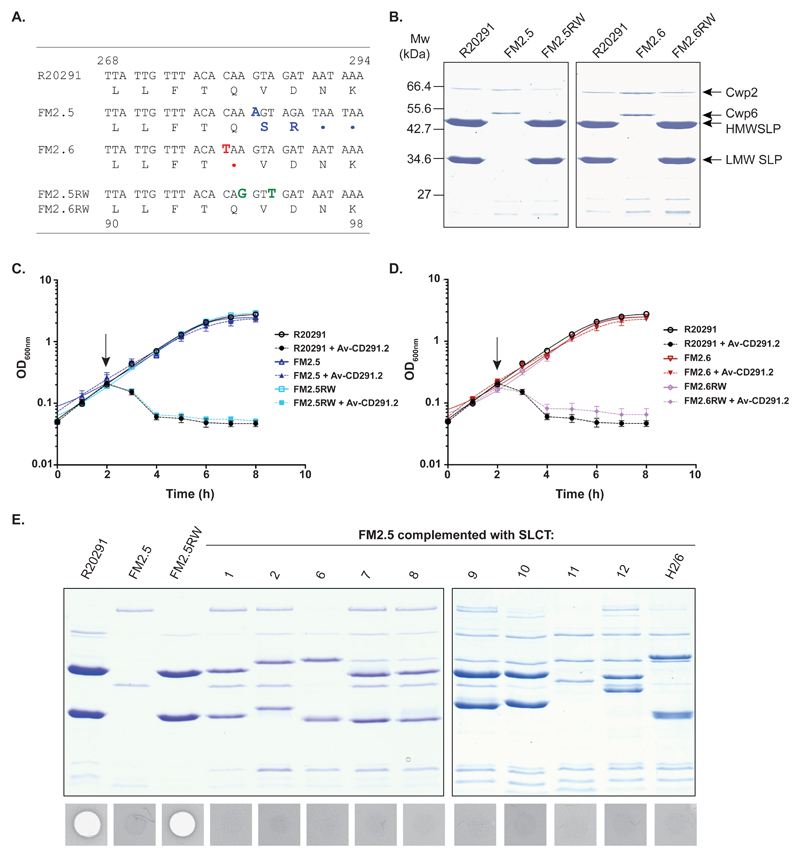
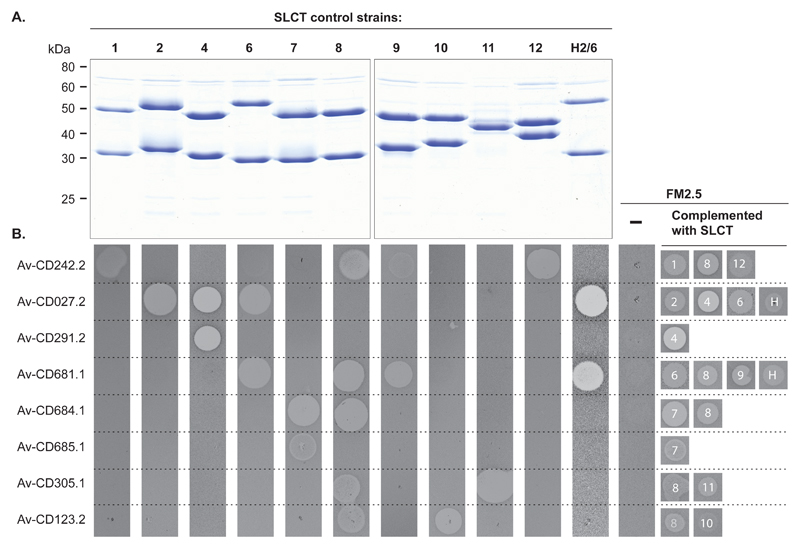
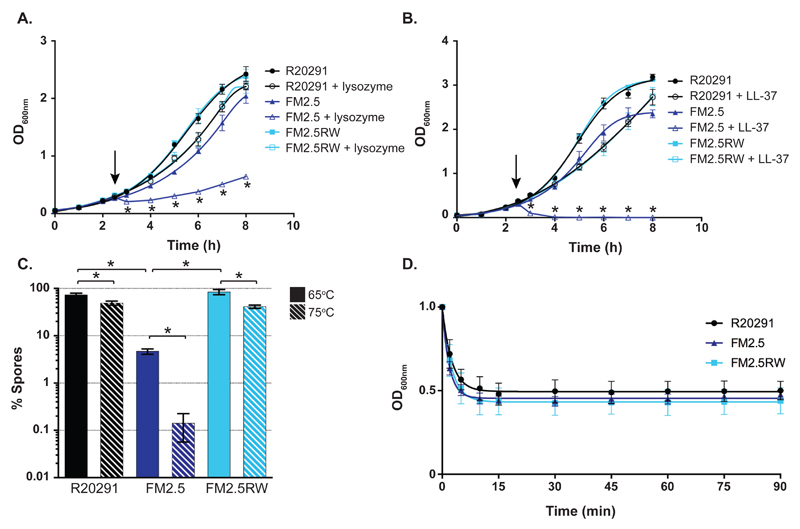
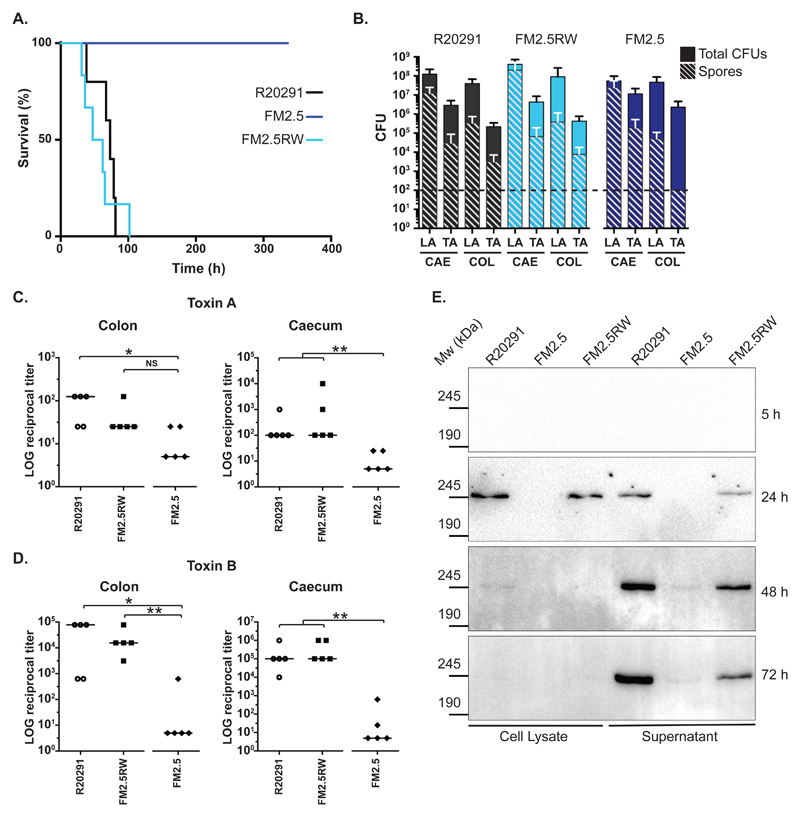
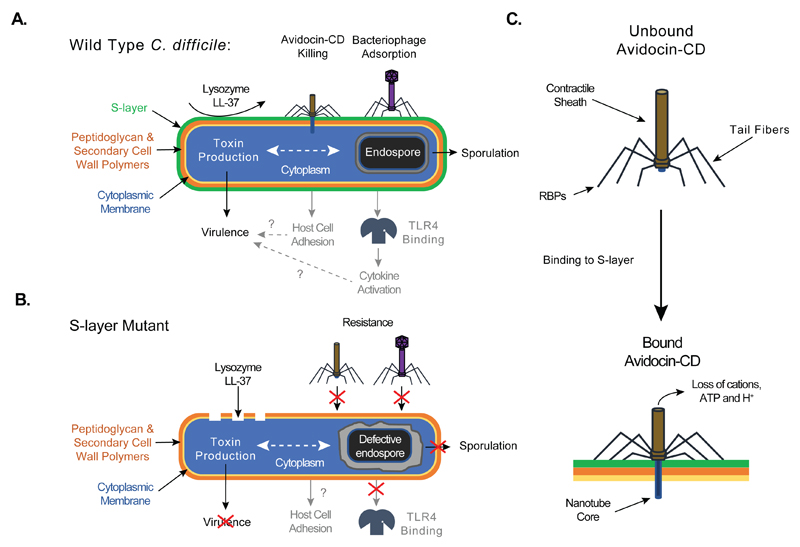
There is a medical need for antibacterial agents that do not damage the resident gut microbiota or promote the spread of antibiotic resistance. We recently described a prototypic precision bactericidal agent, Av-CD291.2, which selectively kills specific Clostridium difficile strains and prevents them from colonizing mice. We have since selected two Av-CD291.2-resistant mutants that have a surface (S)-layer-null phenotype due to distinct point mutations in the slpA gene. Using newly identified bacteriophage receptor binding proteins for targeting, we constructed a panel of Avidocin-CDs that kills diverse C. difficile isolates in an S-layer sequence-dependent manner. In addition to bacteriophage receptor recognition, characterization of the mutants also uncovered important roles for S-layer protein A (SlpA) in sporulation, resistance to innate immunity effectors, and toxin production. Surprisingly, S-layer-null mutants were found to persist in the hamster gut despite a complete attenuation of virulence. These findings suggest antimicrobials targeting virulence factors dispensable for fitness in the host force pathogens to trade virulence for viability and would have clear clinical advantages should resistance emerge. Given their exquisite specificity for the pathogen, Avidocin-CDs have substantial therapeutic potential for the treatment and prevention of C. difficile infections.
Copyright © 2017 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Conflict of interest statement
D.G., S.L., D.S., and G.R.G. are current or past employees of and own stock in AvidBiotics Corp. R.P.F. received a research grant from AvidBiotics Corp. AvidBiotics Corp. hold the following patents: US8206971 (Modified bacteriocins and methods for their use), US8673291 (Diffocins and methods of use therof), US9115354 (Diffocins and methods of use therof), and EP2576604 (Diffocins and methods of use therof).
Figures
References
-
- Rasko DA, Sperandio V. Anti-virulence strategies to combat bacteria-mediated disease. Nat Rev Drug Discov. 2010;9:117–128. - PubMed
-
- Allen RC, Popat R, Diggle SP, Brown SP. Targeting virulence: can we make evolution-proof drugs? Nat Rev Microbiol. 2014;12:300–308. - PubMed
-
- ANTIBIOTIC RESISTANCE THREATS in the United States, 2013, Threat Report 2013. Centers for Disease Control and Prevention; Atlanta: 2013.
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
