

Enantiomeric barbiturates bind distinct inter- and intrasubunit binding sites in a nicotinic acetylcholine receptor (nAChR)
- PMID: 28878016
- PMCID: PMC5655505
- DOI: 10.1074/jbc.M117.808592
Enantiomeric barbiturates bind distinct inter- and intrasubunit binding sites in a nicotinic acetylcholine receptor (nAChR)
Abstract
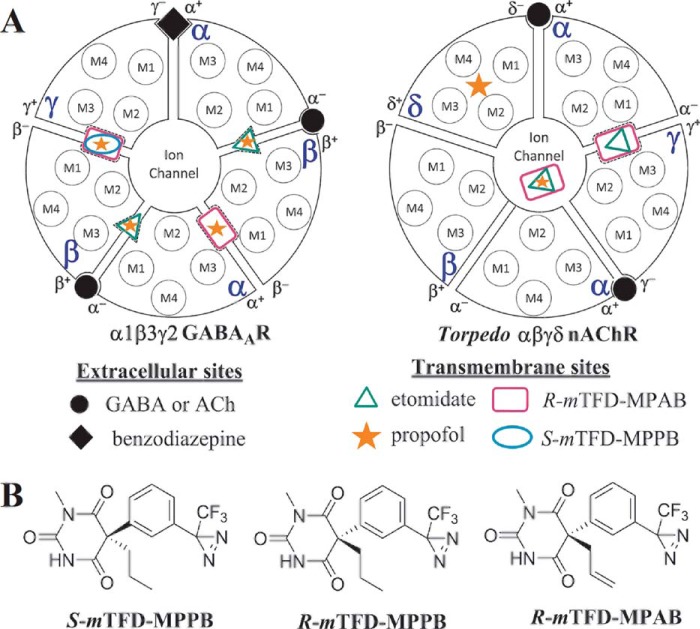
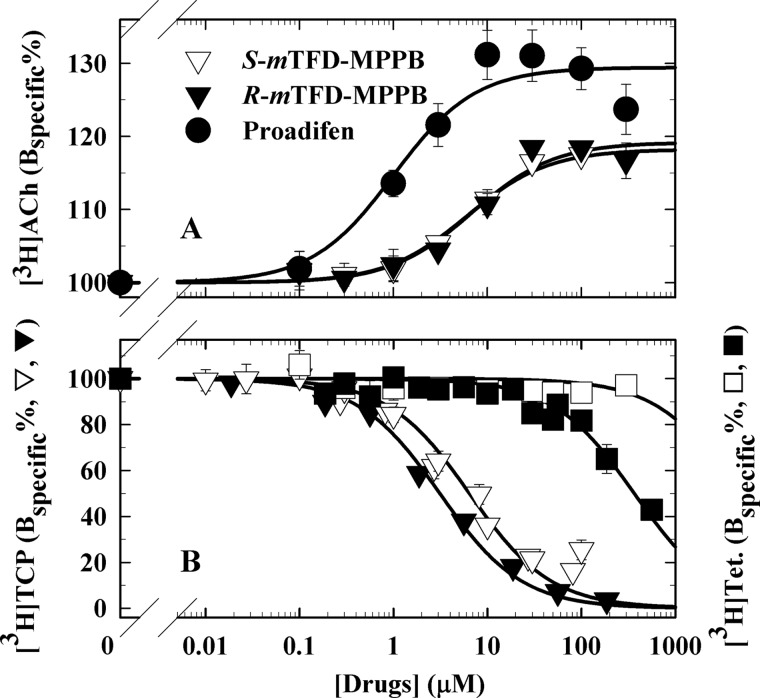
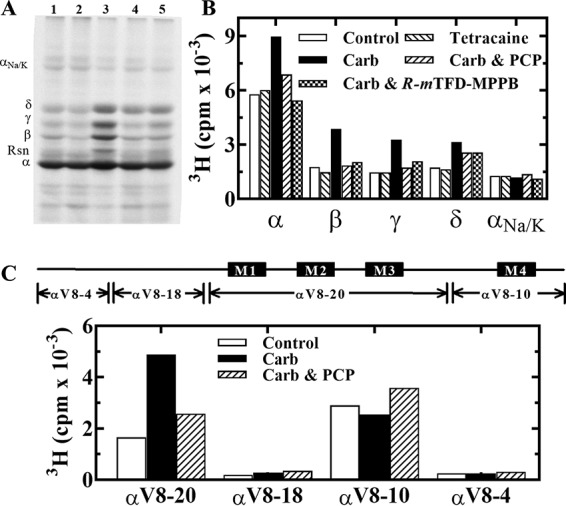
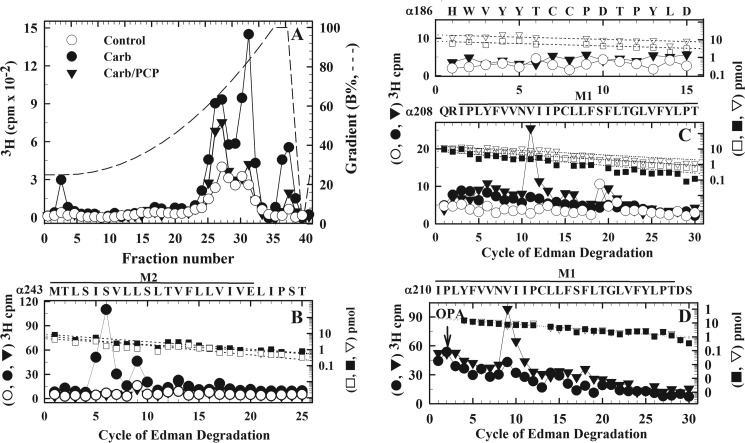
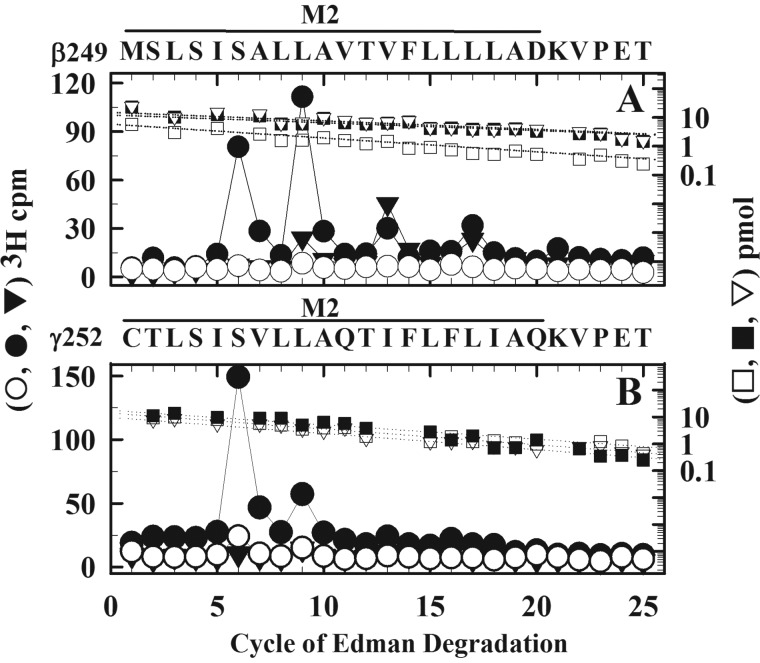
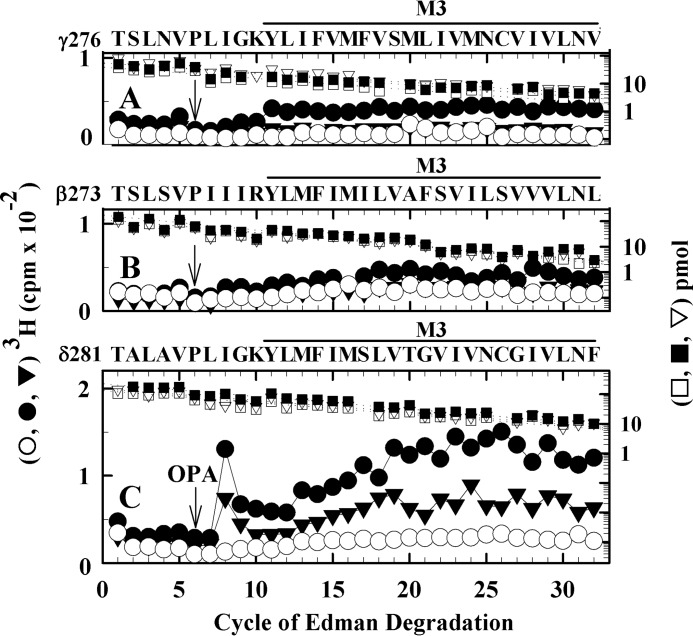
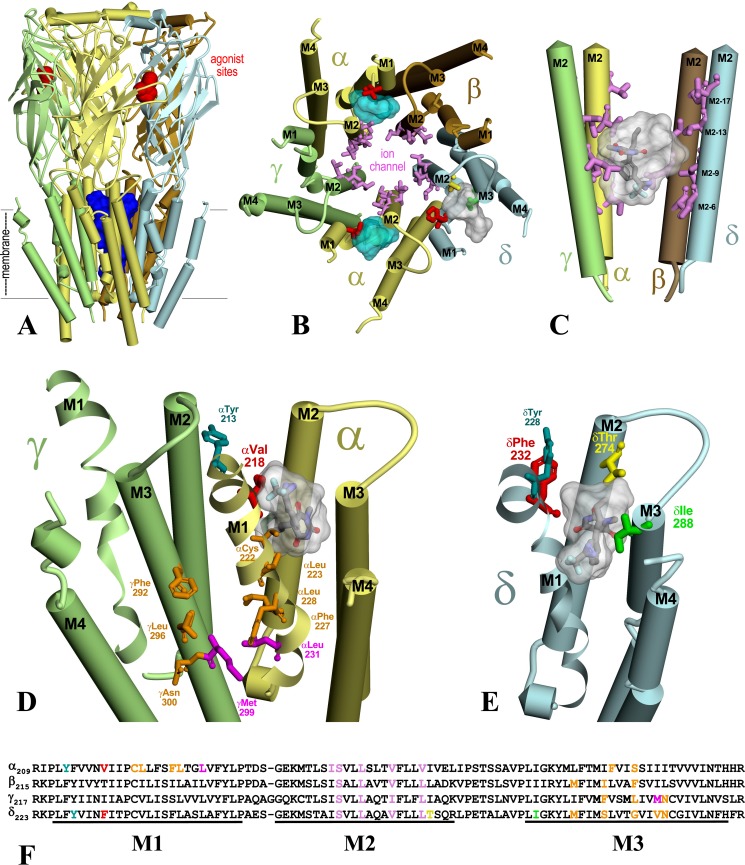
Nicotinic acetylcholine receptors (nAChRs) and γ-aminobutyric acid type A receptors (GABAARs) are members of the pentameric ligand-gated ion channel superfamily. Drugs acting as positive allosteric modulators of muscle-type α2βγδ nAChRs, of use in treatment of neuromuscular disorders, have been hard to identify. However, identification of nAChR allosteric modulator binding sites has been facilitated by using drugs developed as photoreactive GABAAR modulators. Recently, R-1-methyl-5-allyl-5-(m-trifluoromethyl-diazirinylphenyl) barbituric acid (R-mTFD-MPAB), an anesthetic and GABAAR potentiator, has been shown to inhibit Torpedo α2βγδ nAChRs, binding in the ion channel and to a γ+-α- subunit interface site similar to its GABAAR intersubunit binding site. In contrast, S-1-methyl-5-propyl-5-(m-trifluoromethyl-diazirinylphenyl) barbituric acid (S-mTFD-MPPB) acts as a convulsant and GABAAR inhibitor. Photolabeling studies established that S-mTFD-MPPB binds to the same GABAAR intersubunit binding site as R-mTFD-MPAB, but with negative rather than positive energetic coupling to GABA binding. We now show that S-mTFD-MPPB binds with the same state (agonist) dependence as R-mTFD-MPAB within the nAChR ion channel, but it does not bind to the intersubunit binding site. Rather, S-mTFD-MPPB binds to intrasubunit sites within the α and δ subunits, photolabeling αVal-218 (αM1), δPhe-232 (δM1), δThr-274 (δM2), and δIle-288 (δM3). Propofol, a general anesthetic that binds to GABAAR intersubunit sites, inhibited [3H]S-mTFD-MPPB photolabeling of these nAChR intrasubunit binding sites. These results demonstrate that in an nAChR, the subtle difference in structure between S-mTFD-MPPB and R-mTFD-MPAB (chirality; 5-propyl versus 5-allyl) determines selectivity for intra- versus intersubunit sites, in contrast to GABAARs, where this difference affects state dependence of binding to a common site.
Keywords: GABA receptor; allosteric regulation; anesthetic; nicotinic acetylcholine receptors (nAChR); photoaffinity labeling.
© 2017 by The American Society for Biochemistry and Molecular Biology, Inc.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures



References
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
