

Optimum Temperatures for Net Primary Productivity of Three Tropical Seagrass Species
- PMID: 28878790
- PMCID: PMC5572403
- DOI: 10.3389/fpls.2017.01446
Optimum Temperatures for Net Primary Productivity of Three Tropical Seagrass Species
Abstract
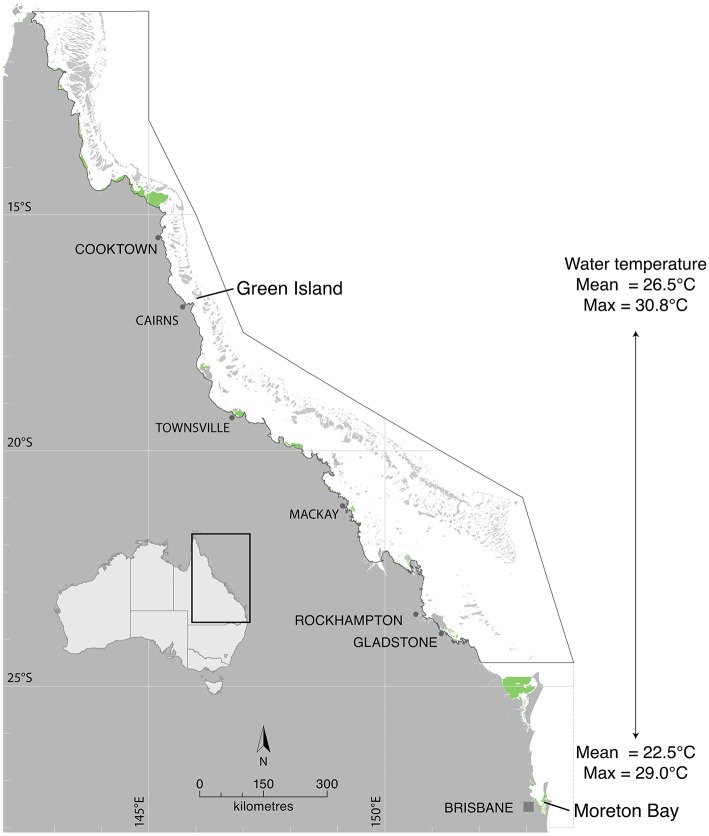
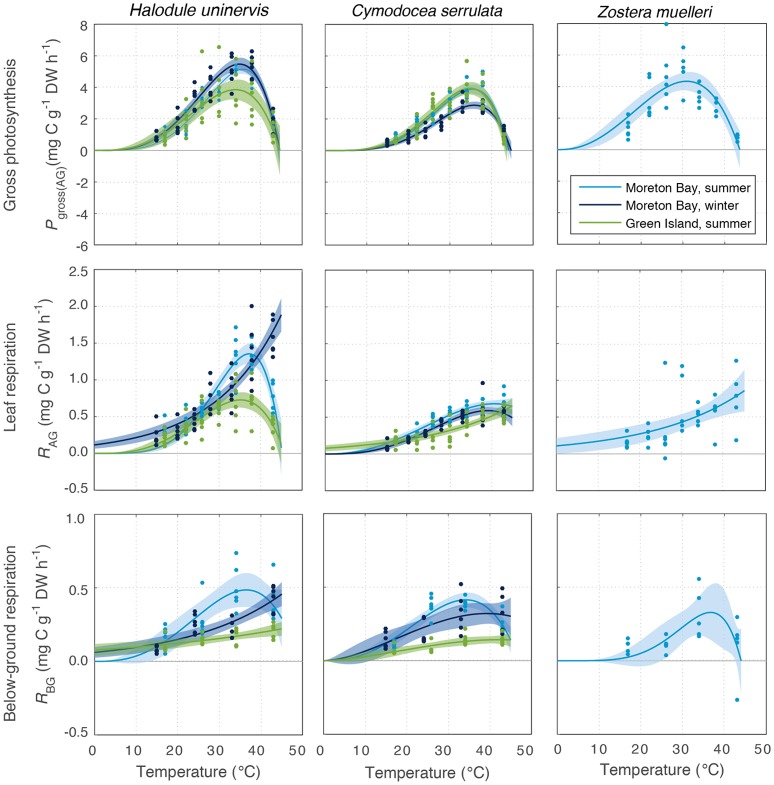
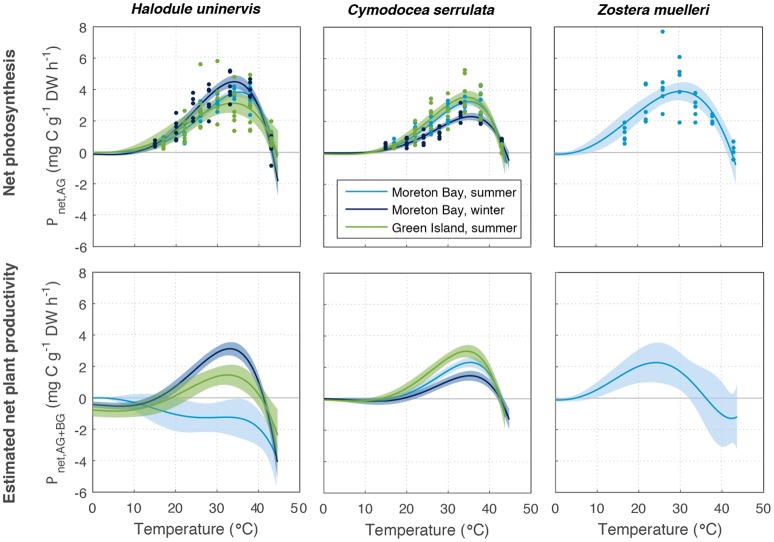
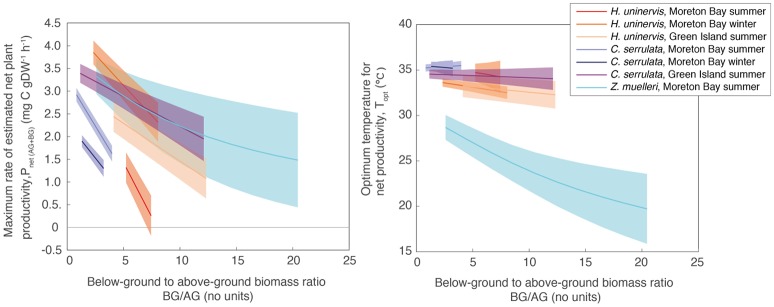
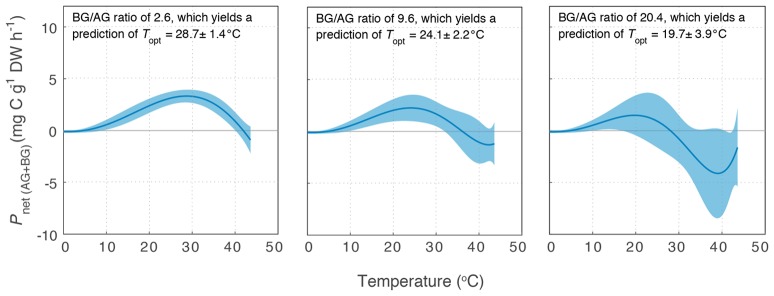
Rising sea water temperature will play a significant role in responses of the world's seagrass meadows to climate change. In this study, we investigated seasonal and latitudinal variation (spanning more than 1,500 km) in seagrass productivity, and the optimum temperatures at which maximum photosynthesis and net productivity (for the leaf and the whole plant) occurs, for three seagrass species (Cymodocea serrulata, Halodule uninervis, and Zostera muelleri). To obtain whole plant net production, photosynthesis, and respiration rates of leaves and the root/rhizome complex were measured using oxygen-sensitive optodes in closed incubation chambers at temperatures ranging from 15 to 43°C. The temperature-dependence of photosynthesis and respiration was fitted to empirical models to obtain maximum metabolic rates and thermal optima. The thermal optimum (Topt) for gross photosynthesis of Z. muelleri, which is more commonly distributed in sub-tropical to temperate regions, was 31°C. The Topt for photosynthesis of the tropical species, H. uninervis and C. serrulata, was considerably higher (35°C on average). This suggests that seagrass species are adapted to water temperature within their distributional range; however, when comparing among latitudes and seasons, thermal optima within a species showed limited acclimation to ambient water temperature (Topt varied by 1°C in C. serrulata and 2°C in H. uninervis, and the variation did not follow changes in ambient water temperature). The Topt for gross photosynthesis were higher than Topt calculated from plant net productivity, which includes above- and below-ground respiration for Z. muelleri (24°C) and H. uninervis (33°C), but remained unchanged at 35°C in C. serrulata. Both estimated plant net productivity and Topt are sensitive to the proportion of below-ground biomass, highlighting the need for consideration of below- to above-ground biomass ratios when applying thermal optima to other meadows. The thermal optimum for plant net productivity was lower than ambient summer water temperature in Z. muelleri, indicating likely contemporary heat stress. In contrast, thermal optima of H. uninervis and C. serrulata exceeded ambient water temperature. This study found limited capacity to acclimate: thus the thermal optima can forewarn of both the present and future vulnerability to ocean warming during periods of elevated water temperature.
Keywords: Cymodocea serrulata; Halodule uninervis; Zostera muelleri; climate change; net primary productivity; sea temperature; thermal stress; tropical seagrass.
Figures
References
-
- Anthony K. R. N., Hoogenboom M. O., Maynard J. A., Grottoli A. G., Middlebrook R. (2009). Energetics approach to predicting mortality risk from environmental stress: a case study of coral bleaching. Funct. Ecol. 23, 539–550. 10.1111/j.1365-2435.2008.01531.x - DOI
-
- Baird M. E., Adams M. P., Babcock R. C., Oubelkheir K., Mongin M., Wild-Allen K. A., et al. (2016). A biophysical representation of seagrass growth for application in a complex shallow-water biogeochemical model. Ecol. Modell. 325, 13–27. 10.1016/j.ecolmodel.2015.12.011 - DOI
LinkOut - more resources
Full Text Sources
Other Literature Sources