

Genome sequence of the sulfur-oxidizing Bathymodiolus thermophilus gill endosymbiont
- PMID: 28878861
- PMCID: PMC5581435
- DOI: 10.1186/s40793-017-0266-y
Genome sequence of the sulfur-oxidizing Bathymodiolus thermophilus gill endosymbiont
Abstract
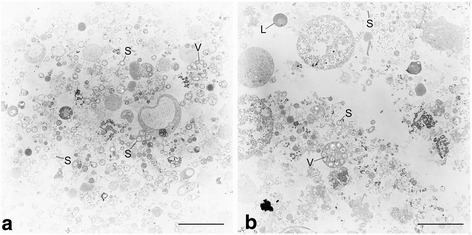
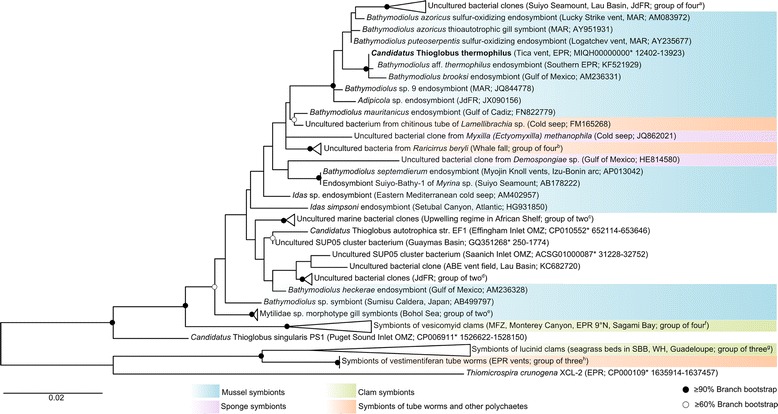
Bathymodiolus thermophilus, a mytilid mussel inhabiting the deep-sea hydrothermal vents of the East Pacific Rise, lives in symbiosis with chemosynthetic Gammaproteobacteria within its gills. The intracellular symbiont population synthesizes nutrients for the bivalve host using the reduced sulfur compounds emanating from the vents as energy source. As the symbiont is uncultured, comprehensive and detailed insights into its metabolism and its interactions with the host can only be obtained from culture-independent approaches such as genomics and proteomics. In this study, we report the first draft genome sequence of the sulfur-oxidizing symbiont of B. thermophilus, here tentatively named Candidatus Thioglobus thermophilus. The draft genome (3.1 Mb) harbors 3045 protein-coding genes. It revealed pathways for the use of sulfide and thiosulfate as energy sources and encodes the Calvin-Benson-Bassham cycle for CO2 fixation. Enzymes required for the synthesis of the tricarboxylic acid cycle intermediates oxaloacetate and succinate were absent, suggesting that these intermediates may be substituted by metabolites from external sources. We also detected a repertoire of genes associated with cell surface adhesion, bacteriotoxicity and phage immunity, which may perform symbiosis-specific roles in the B. thermophilus symbiosis.
Keywords: Autotrophy; Hydrothermal vents; Marine invertebrate symbiosis; Thiotrophy; Uncultured endosymbiont.
Conflict of interest statement
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures
References
-
- Karl DM, Wirsen CO, Jannasch HW. Deep-sea primary production at the galapagos hydrothermal vents. Science. 1980;207(4437):1345–1347. doi: 10.1126/science.207.4437.1345. - DOI
-
- Felbeck H, Somero GN. Primary production in deep-sea hydrothermal vent organisms: roles of sulfide-oxidizing bacteria. Trends Biochem Sci. 1982;7(6):201–204. doi: 10.1016/0968-0004(82)90088-3. - DOI
-
- Cavanaugh CM. Symbiotic chemoautotrophic bacteria in marine invertebrates from sulfide-rich habitats. Nature. 1983;302(5903):58–61. doi: 10.1038/302058a0. - DOI
-
- Van Dover CL. The ecology of deep-sea hydrothermal vents. 1. New Jersey: Princeton University Press; 2000.
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
