

ARF6 mediates nephrin tyrosine phosphorylation-induced podocyte cellular dynamics
- PMID: 28880939
- PMCID: PMC5589247
- DOI: 10.1371/journal.pone.0184575
ARF6 mediates nephrin tyrosine phosphorylation-induced podocyte cellular dynamics
Abstract
ADP-ribosylation factor 6 (ARF6) is a small GTPase necessary for regulating cellular structure, motility, and vesicle trafficking. In several cellular systems, ARF6 was shown to regulate actin dynamics in coordination with Rac1, a Rho small GTPase. We examined the function of ARF6 in the kidney podocyte because Rac1 was implicated in kidney diseases involving this cell. We found that ARF6 expression was enriched in human podocytes and that it modulated podocyte cytoskeletal dynamics through a functional interaction with nephrin, an intercellular junction protein necessary for podocyte injury-induced signaling requiring activation by tyrosine phosphorylation of its cytoplasmic domain. ARF6 was necessary for nephrin activation-induced ruffling and focal adhesion turnover, possibly by altering Rac1 activity. In podocyte-specific Arf6 (ARF6_PodKO) knockout mice, ARF6 deficiency did not result in a spontaneous kidney developmental phenotype or proteinuria after aging. However, ARF6_PodKO mice exhibited distinct phenotypes in two in vivo glomerular injury models. In the protamine sulfate perfusion model, which induced acute podocyte effacement, ARF6_PodKO mice were protected from podocyte effacement. In the nephrotoxic serum nephritis model, which induced immune-complex mediated injury, ARF6_PodKO mice exhibited aggravated proteinuria. Together, these observations suggest that while ARF6 is necessary for nephrin tyrosine phosphorylation-induced cytoskeletal dynamics in cultured podocytes, ARF6 has pleotropic podocyte roles in vivo, where glomerular injury-specific mechanisms might activate distinct signaling pathways that dictate whether ARF6 activity is beneficial or deleterious for maintaining the integrity of the glomerular filtration barrier.
Conflict of interest statement
Figures
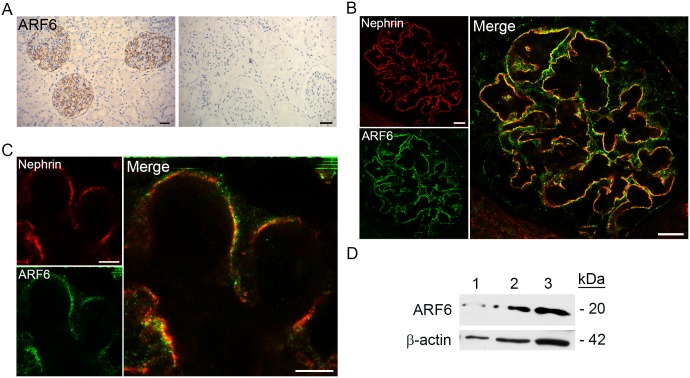
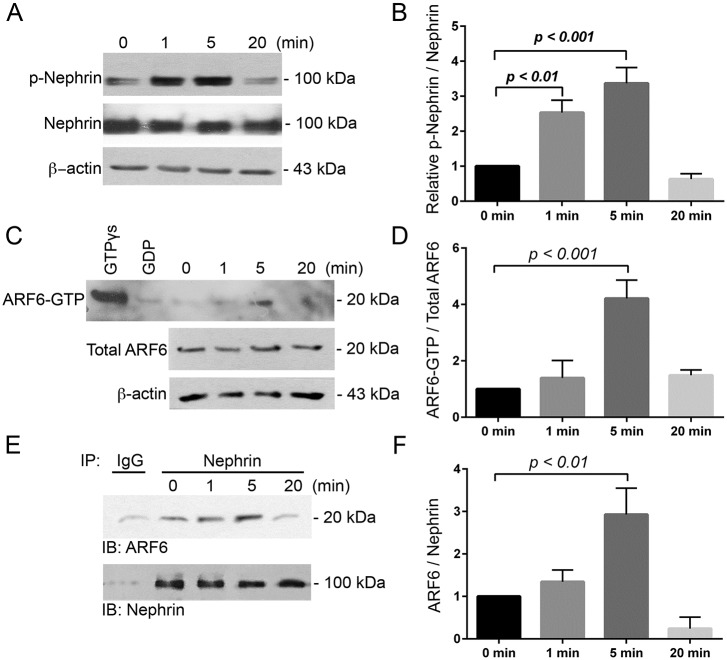
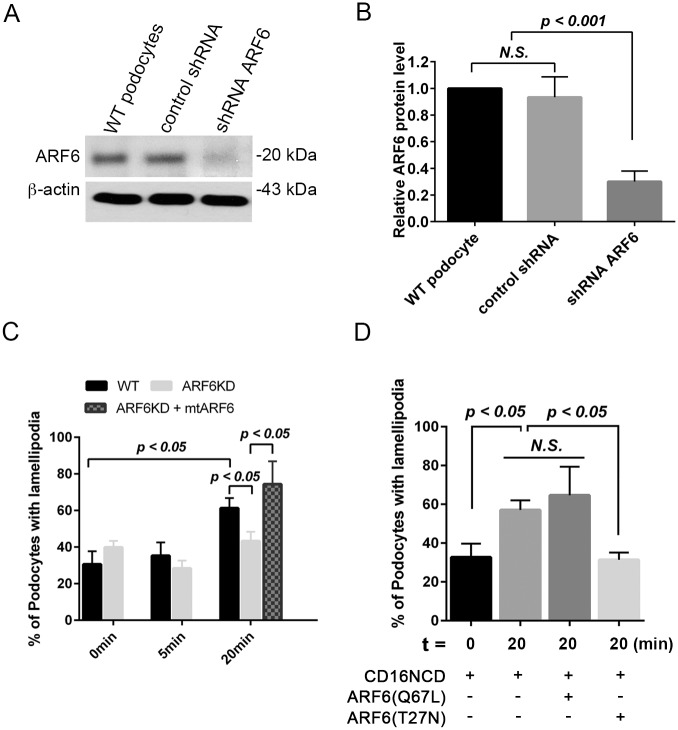
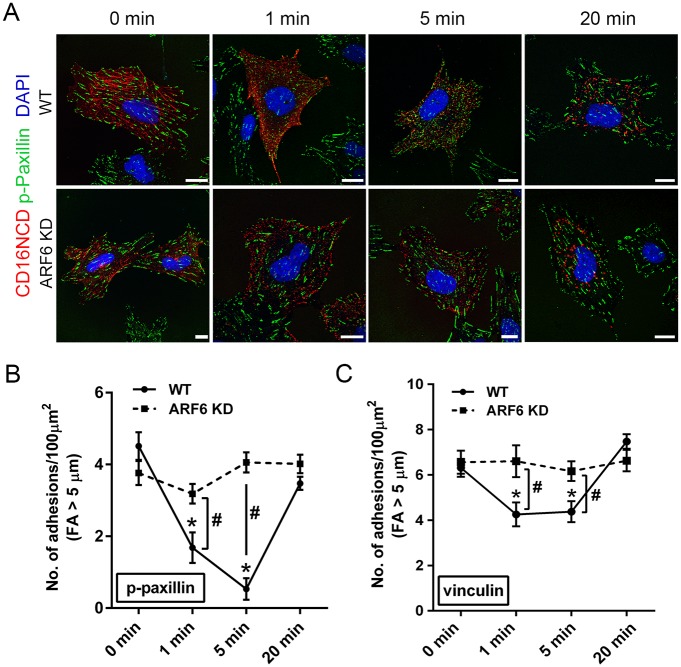
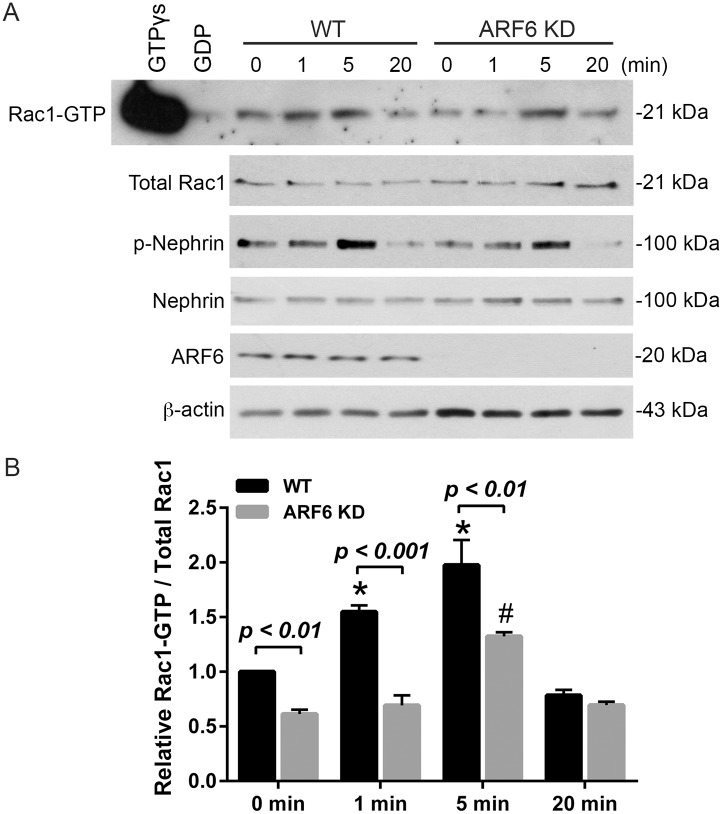
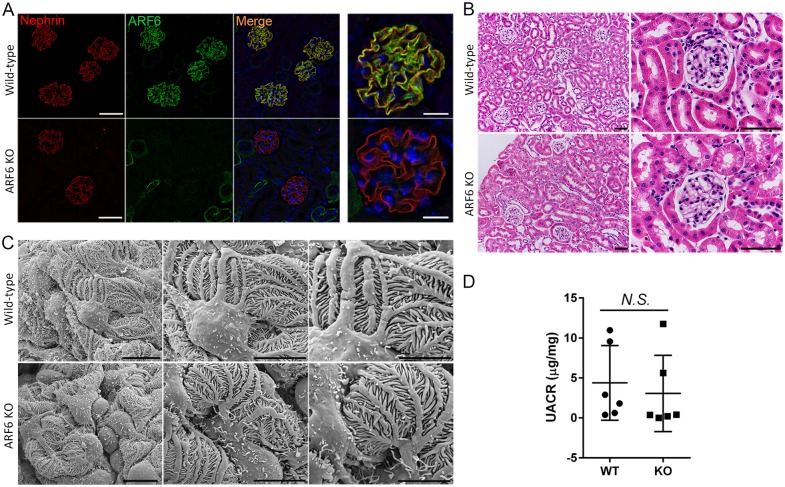
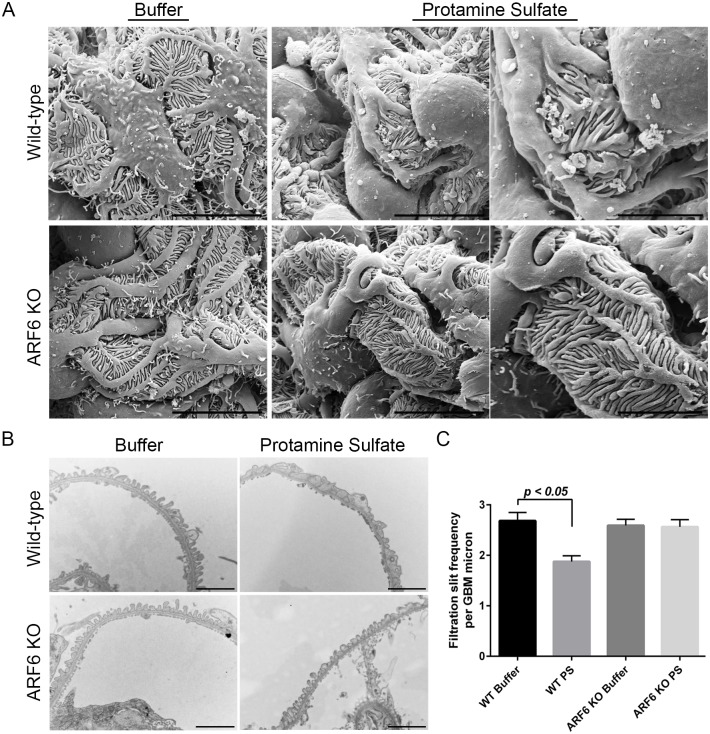
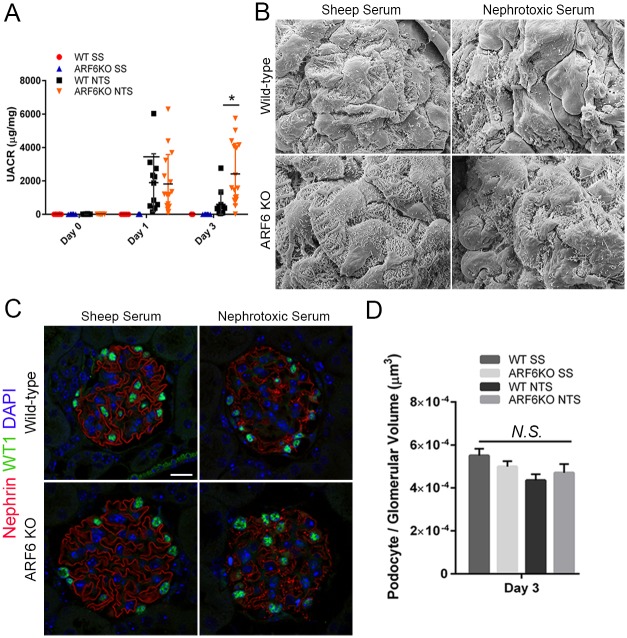
References
-
- Mundel P, Shankland SJ. Podocyte biology and response to injury. Journal of the American Society of Nephrology: JASN. 2002;13(12):3005–15. - PubMed
-
- Oh J, Reiser J, Mundel P. Dynamic (re)organization of the podocyte actin cytoskeleton in the nephrotic syndrome. Pediatric nephrology (Berlin, Germany). 2004;19(2):130–7. - PubMed
-
- Ichikawa I, Ma J, Motojima M, Matsusaka T. Podocyte damage damages podocytes: autonomous vicious cycle that drives local spread of glomerular sclerosis. Current opinion in nephrology and hypertension. 2005;14(3):205–10. - PubMed
-
- Wharram BL, Goyal M, Wiggins JE, Sanden SK, Hussain S, Filipiak WE, et al. Podocyte depletion causes glomerulosclerosis: diphtheria toxin-induced podocyte depletion in rats expressing human diphtheria toxin receptor transgene. Journal of the American Society of Nephrology: JASN. 2005;16(10):2941–52. doi: 10.1681/ASN.2005010055 - DOI - PubMed
-
- Patrakka J, Tryggvason K. New insights into the role of podocytes in proteinuria. Nature reviews Nephrology. 2009;5(8):463–8. doi: 10.1038/nrneph.2009.108 - DOI - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
