

C/EBP-δ positively regulates MDSC expansion and endothelial VEGFR2 expression in tumor development
- PMID: 28881585
- PMCID: PMC5584171
- DOI: 10.18632/oncotarget.16410
C/EBP-δ positively regulates MDSC expansion and endothelial VEGFR2 expression in tumor development
Abstract
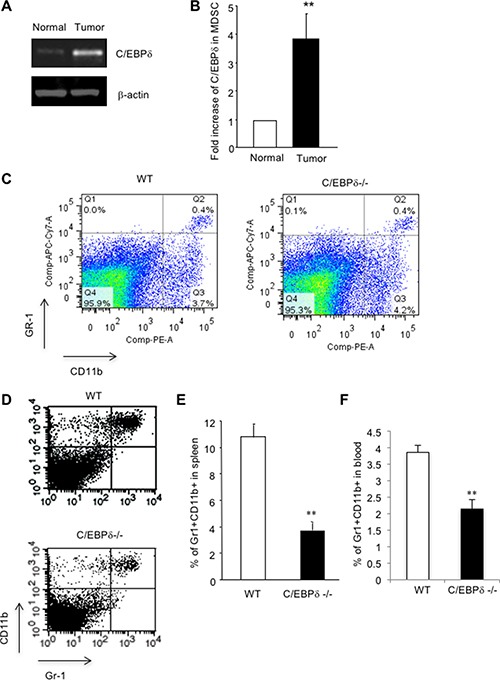
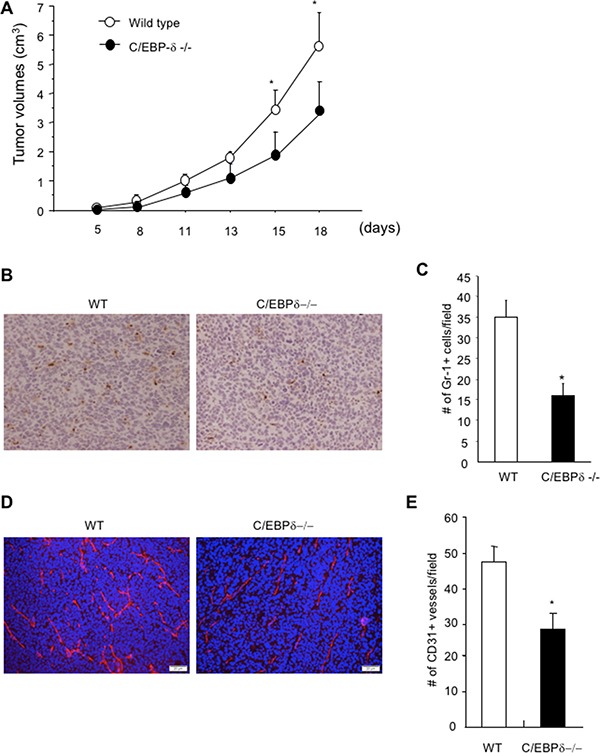
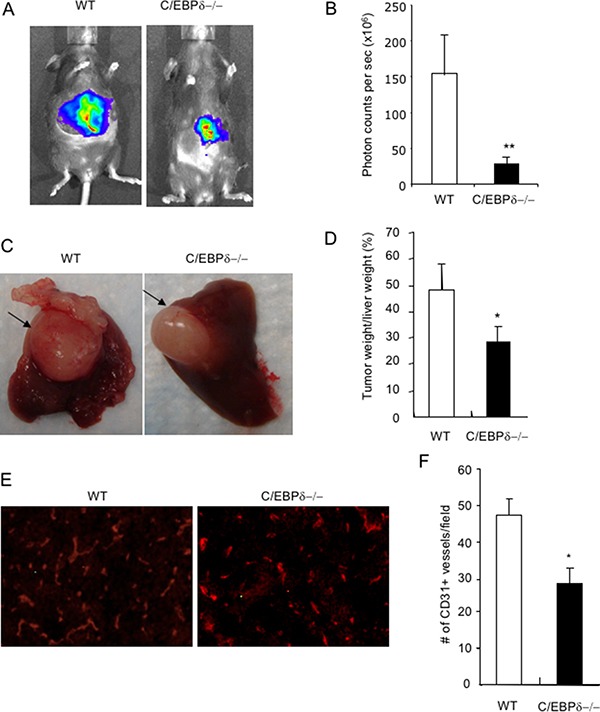
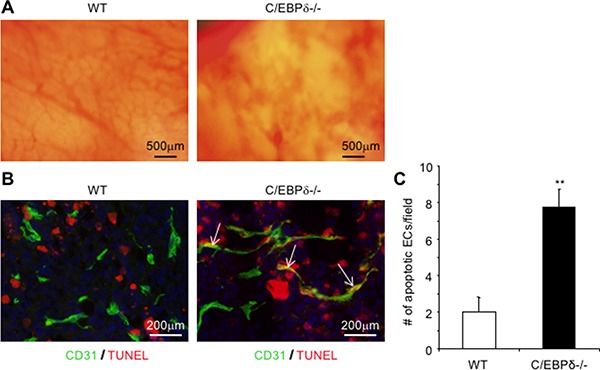
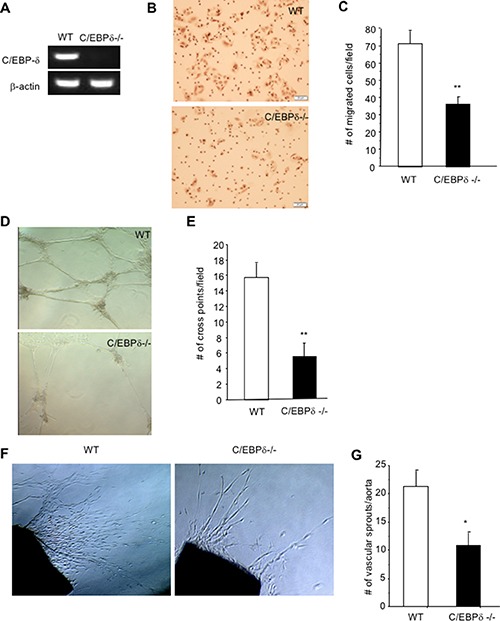
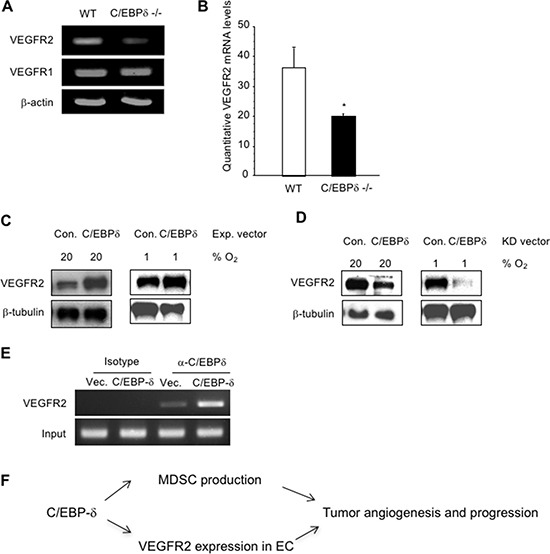
Vascular endothelial cells and Gr-1+CD11b+ myeloid derived suppressor cells (MDSCs) are two important components that constitute the tumor microenvironment. Targeting these cells offers the potential to halt tumor growth. In this study, we report a common mediator in C/EBP-δ that regulates both components and aids in tumor development. C/EBP-δ is elevated in tumor derived MDSCs. Interestingly, genetic deletion of C/EBP-δ in mice significantly impaired MDSC expansion in response to tumor progression, but it had no effect on Gr-1+CD11b+ cell production in normal development. It suggests a specific role of C/EBP-δ in emergency myelopoiesis under tumor conditions. Consistent with the pro tumor functions of MDSCs, loss of C/EBP-δ resulted in reduced tumor angiogenesis and tumor growth. Moreover, we found expression of C/EBP-δ in vascular endothelial cells. C/EBP-δ regulated cell motility, endothelial network formation and vascular sprouting. Notably, inactivation of C/EBP-δ in endothelial cells specifically inhibited the expression of VEGFR2 but not VEGFR1. Ectopic expression of C/EBP-δ increased and knockdown of the gene decreased VEGFR2 expression. C/EBP-δ is recruited to the promoter region of VEGFR2, indicative of transcriptional regulation. Collectively, this study has identified a positive mediator in C/EBP-δ, which regulates tumor induced MDSC expansion and VEGFR2 expression in endothelium. Considering the importance of MDSCs and endothelial cells in tumor progression, targeting C/EBP-δ may provide an interesting means for cancer therapy, killing two birds with one stone.
Keywords: C/EBP-δ; MDSCs; VEGFR2; angiogenesis; cancer.
Conflict of interest statement
CONFLICTS OF INTEREST The authors declare no conflicts of interest.
Figures
References
-
- Bjerregaard MD, Jurlander J, Klausen P, Borregaard N, Cowland JB. The in vivo profile of transcription factors during neutrophil differentiation in human bone marrow. Blood. 2003;101:4322–4332. - PubMed
-
- Chang W, Parra M, Centrella M, McCarthy TL. Interactions between CCAAT enhancer binding protein delta and estrogen receptor alpha control insulin-like growth factor I (igf1) and estrogen receptor-dependent gene expression in osteoblasts. Gene. 2005;345:225–235. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
