

Kinetic signatures of myosin-5B, the motor involved in microvillus inclusion disease
- PMID: 28882893
- PMCID: PMC5672057
- DOI: 10.1074/jbc.M117.801456
Kinetic signatures of myosin-5B, the motor involved in microvillus inclusion disease
Abstract
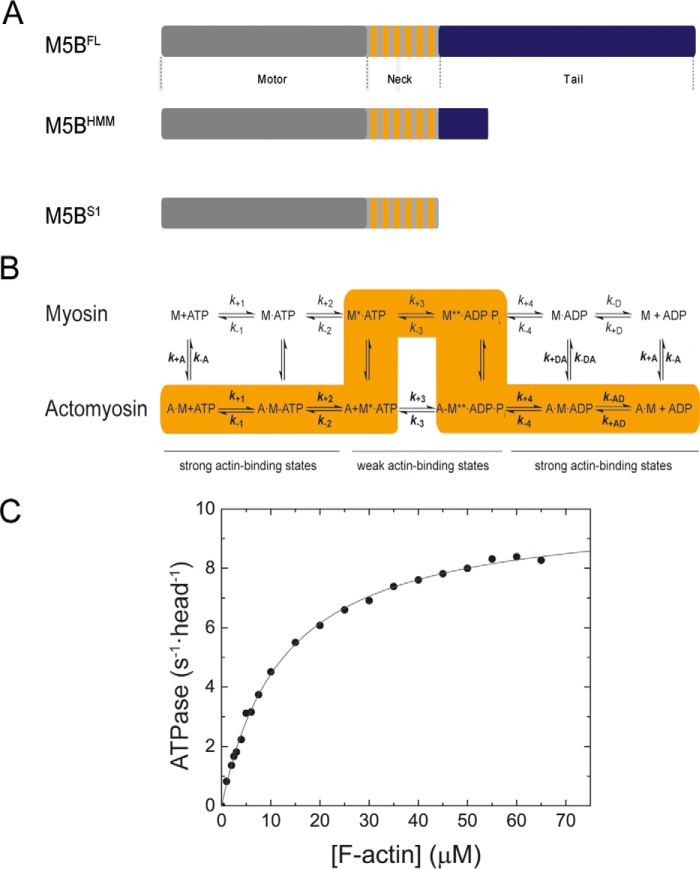
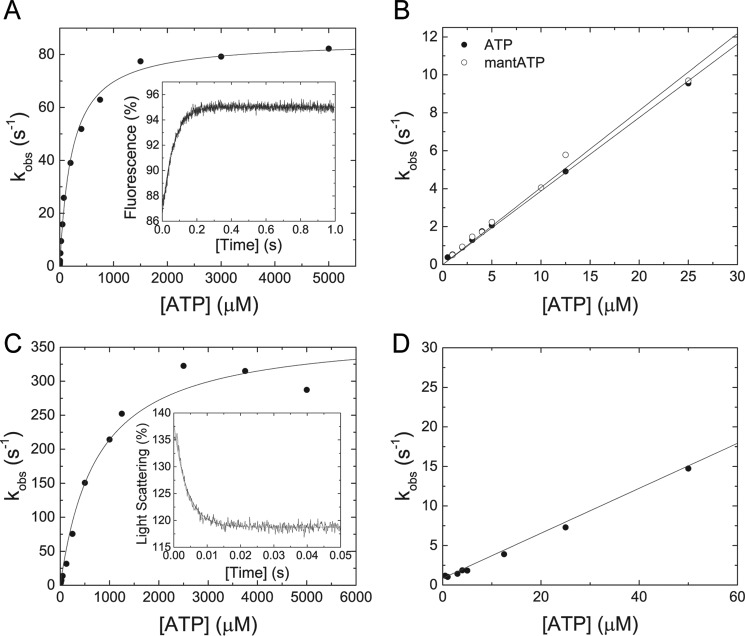
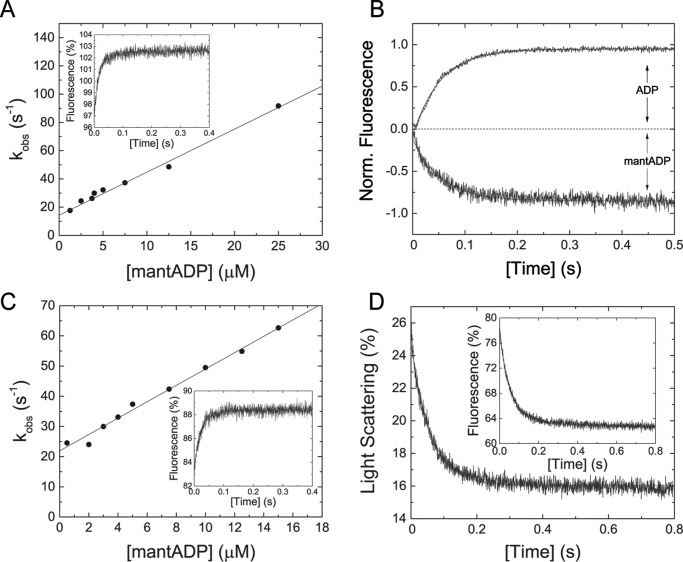
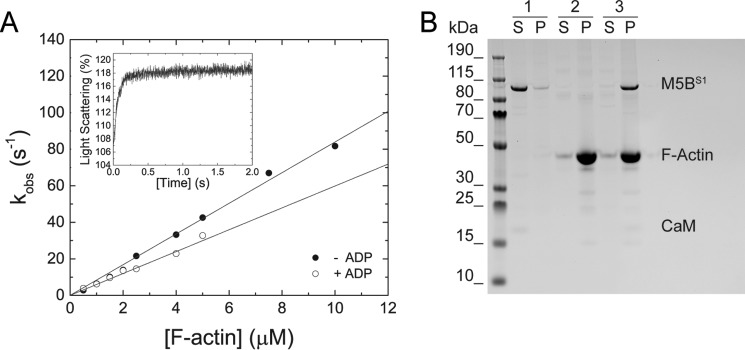
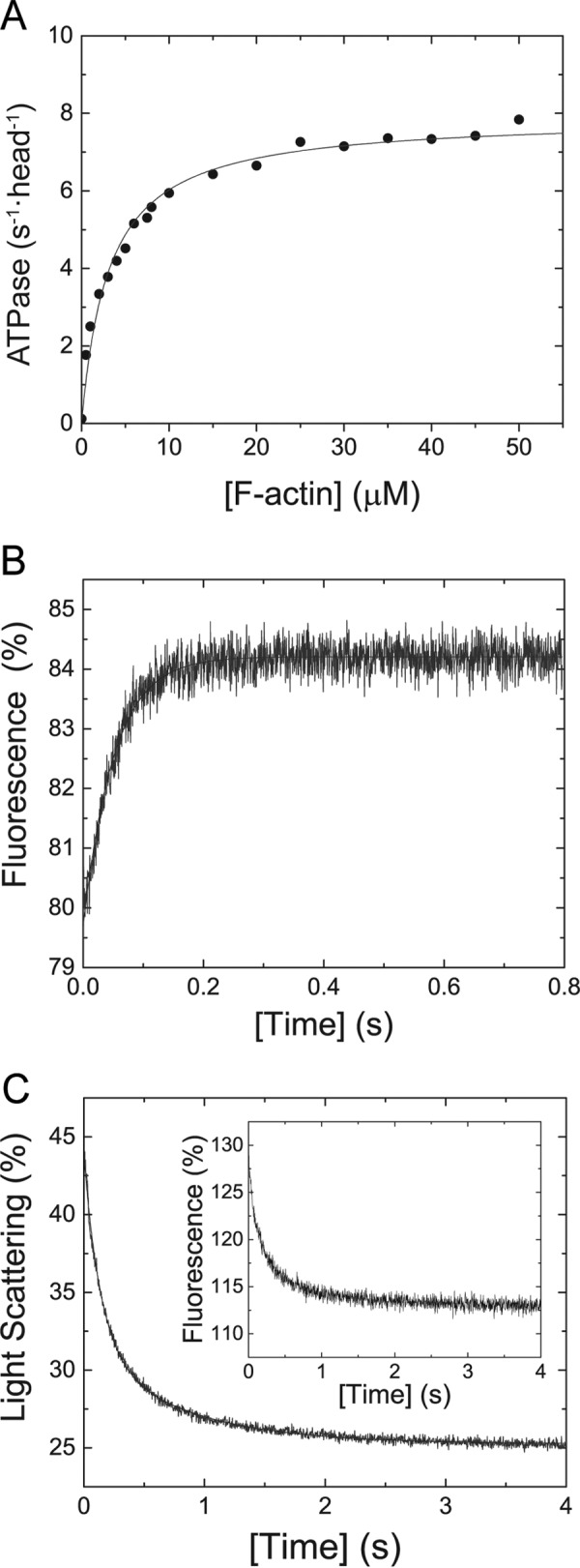
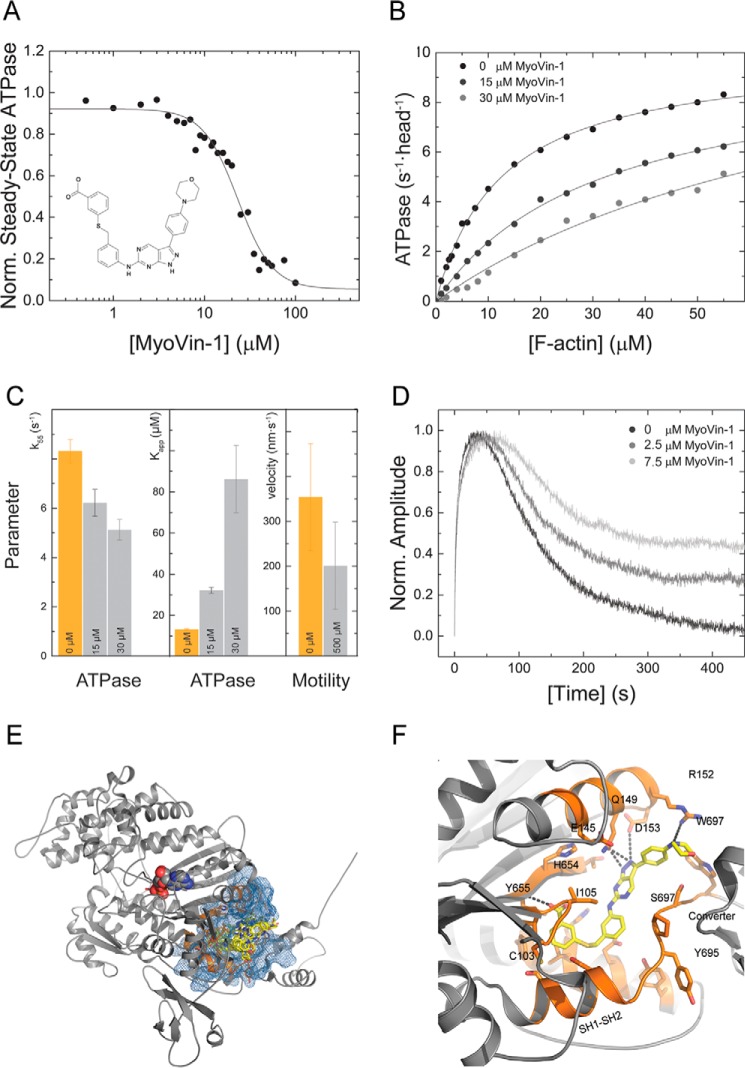
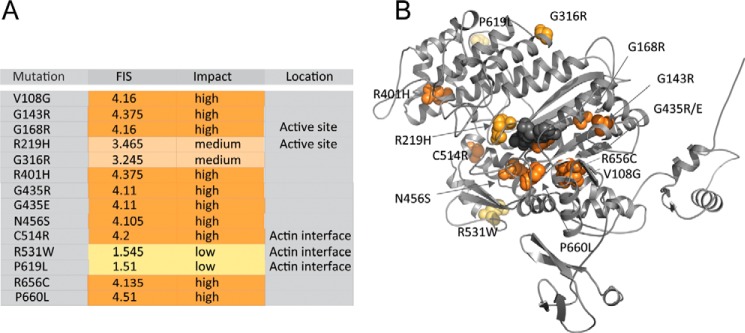
Myosin-5B is a ubiquitous molecular motor that transports cargo vesicles of the endomembrane system in intracellular recycling pathways. Myosin-5B malfunction causes the congenital enteropathy microvillus inclusion disease, underlining its importance in cellular homeostasis. Here we describe the interaction of myosin-5B with F-actin, nucleotides, and the pyrazolopyrimidine compound myoVin-1. We show that single-headed myosin-5B is an intermediate duty ratio motor with a kinetic ATPase cycle that is rate-limited by the release of phosphate. The presence of a second head generates strain and gating in the myosin-5B dimer that alters the kinetic signature by reducing the actin-activated ADP release rate to become rate-limiting. This kinetic transition into a high-duty ratio motor is a prerequisite for the proposed transport function of myosin-5B in cellular recycling pathways. Moreover, we show that the small molecule compound myoVin-1 inhibits the enzymatic and functional activity of myosin-5B in vitro Partial inhibition of the actin-activated steady-state ATPase activity and sliding velocity suggests that caution should be used when probing the effect of myoVin-1 on myosin-5-dependent transport processes in cells.
Keywords: ATPase; actin; inhibitor; kinetics; myoVin-1; myosin.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
Similar articles
-
Microvillus inclusion disease: loss of Myosin vb disrupts intracellular traffic and cell polarity.Traffic. 2014 Jan;15(1):22-42. doi: 10.1111/tra.12131. Epub 2013 Nov 19. Traffic. 2014. PMID: 24138727
-
An overview and online registry of microvillus inclusion disease patients and their MYO5B mutations.Hum Mutat. 2013 Dec;34(12):1597-605. doi: 10.1002/humu.22440. Epub 2013 Oct 16. Hum Mutat. 2013. PMID: 24014347 Review.
-
Myosin 5b loss of function leads to defects in polarized signaling: implication for microvillus inclusion disease pathogenesis and treatment.Am J Physiol Gastrointest Liver Physiol. 2014 Nov 15;307(10):G992-G1001. doi: 10.1152/ajpgi.00180.2014. Epub 2014 Sep 25. Am J Physiol Gastrointest Liver Physiol. 2014. PMID: 25258405 Free PMC article.
-
Myosin Vb uncoupling from RAB8A and RAB11A elicits microvillus inclusion disease.J Clin Invest. 2014 Jul;124(7):2947-62. doi: 10.1172/JCI71651. Epub 2014 Jun 2. J Clin Invest. 2014. PMID: 24892806 Free PMC article.
-
Unequal Effects of Myosin 5B Mutations in Liver and Intestine Determine the Clinical Presentation of Low-Gamma-Glutamyltransferase Cholestasis.Hepatology. 2020 Oct;72(4):1461-1468. doi: 10.1002/hep.31430. Hepatology. 2020. PMID: 32583448 Free PMC article. Review. No abstract available.
Cited by
-
A mutation in switch I alters the load-dependent kinetics of myosin Va.Nat Commun. 2023 May 30;14(1):3137. doi: 10.1038/s41467-023-38535-0. Nat Commun. 2023. PMID: 37253724 Free PMC article.
-
Dissecting myosin-5B mechanosensitivity and calcium regulation at the single molecule level.Nat Commun. 2018 Jul 20;9(1):2844. doi: 10.1038/s41467-018-05251-z. Nat Commun. 2018. PMID: 30030431 Free PMC article.
-
Motor properties of Myosin 5c are modulated by tropomyosin isoforms and inhibited by pentabromopseudilin.Front Physiol. 2024 Mar 28;15:1394040. doi: 10.3389/fphys.2024.1394040. eCollection 2024. Front Physiol. 2024. PMID: 38606007 Free PMC article.
-
The dynamics of actin protrusions can be controlled by tip-localized myosin motors.J Biol Chem. 2024 Jan;300(1):105516. doi: 10.1016/j.jbc.2023.105516. Epub 2023 Nov 30. J Biol Chem. 2024. PMID: 38042485 Free PMC article.
-
Roles and regulation of myosin V interaction with cargo.Adv Biol Regul. 2021 Jan;79:100787. doi: 10.1016/j.jbior.2021.100787. Epub 2021 Jan 20. Adv Biol Regul. 2021. PMID: 33541831 Free PMC article. Review.
References
-
- Hammer J. A. 3rd, and Sellers J. R. (2011) Walking to work: roles for class V myosins as cargo transporters. Nat. Rev. Mol. Cell Biol. 13, 13–26 - PubMed
-
- Nascimento A. A., Roland J. T., and Gelfand V. I. (2003) Pigment cells: a model for the study of organelle transport. Annu. Rev. Cell Dev. Biol. 19, 469–491 - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
