

PP2A-B' holoenzyme substrate recognition, regulation and role in cytokinesis
- PMID: 28884018
- PMCID: PMC5586252
- DOI: 10.1038/celldisc.2017.27
PP2A-B' holoenzyme substrate recognition, regulation and role in cytokinesis
Abstract
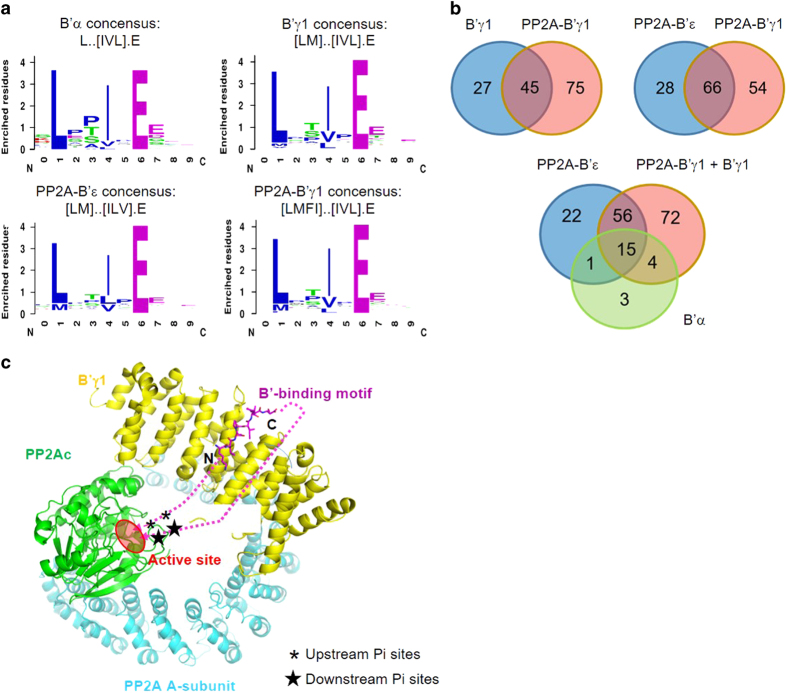
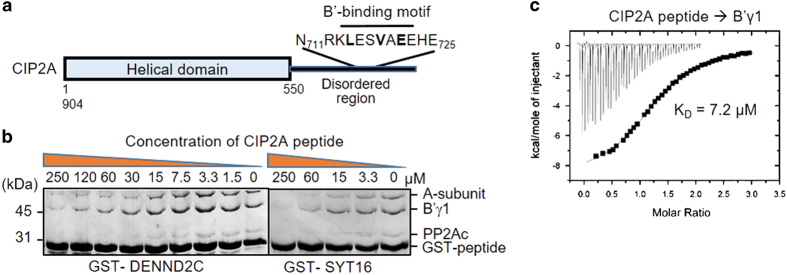
Protein phosphatase 2A (PP2A) is a major Ser/Thr phosphatase; it forms diverse heterotrimeric holoenzymes that counteract kinase actions. Using a peptidome that tiles the disordered regions of the human proteome, we identified proteins containing [LMFI]xx[ILV]xEx motifs that serve as interaction sites for B'-family PP2A regulatory subunits and holoenzymes. The B'-binding motifs have important roles in substrate recognition and in competitive inhibition of substrate binding. With more than 100 novel ligands identified, we confirmed that the recently identified LxxIxEx B'α-binding motifs serve as common binding sites for B' subunits with minor variations, and that S/T phosphorylation or D/E residues at positions 2, 7, 8 and 9 of the motifs reinforce interactions. Hundreds of proteins in the human proteome harbor intrinsic or phosphorylation-responsive B'-interaction motifs, and localize at distinct cellular organelles, such as midbody, predicting kinase-facilitated recruitment of PP2A-B' holoenzymes for tight spatiotemporal control of phosphorylation at mitosis and cytokinesis. Moroever, Polo-like kinase 1-mediated phosphorylation of Cyk4/RACGAP1, a centralspindlin component at the midbody, facilitates binding of both RhoA guanine nucleotide exchange factor (epithelial cell transforming sequence 2 (Ect2)) and PP2A-B' that in turn dephosphorylates Cyk4 and disrupts Ect2 binding. This feedback signaling loop precisely controls RhoA activation and specifies a restricted region for cleavage furrow ingression. Our results provide a framework for further investigation of diverse signaling circuits formed by PP2A-B' holoenzymes in various cellular processes.
Keywords: CIP2A; PP2A-B′ holoenzyme; SLiMs; centrosome; cytokinesis; midbody.
Conflict of interest statement
The authors declare no conflict of interest.
Figures



Similar articles
-
Reduction of protein phosphatase 2A (PP2A) complexity reveals cellular functions and dephosphorylation motifs of the PP2A/B'δ holoenzyme.J Biol Chem. 2020 Apr 24;295(17):5654-5668. doi: 10.1074/jbc.RA119.011270. Epub 2020 Mar 10. J Biol Chem. 2020. PMID: 32156701 Free PMC article.
-
Developmental expression and co-localization of cyclin G1 and the B' subunits of protein phosphatase 2a in neurons.Brain Res Mol Brain Res. 1999 Jan 22;64(1):1-10. doi: 10.1016/s0169-328x(98)00283-6. Brain Res Mol Brain Res. 1999. PMID: 9889295
-
The Brassinosteroid-Activated BRI1 Receptor Kinase Is Switched off by Dephosphorylation Mediated by Cytoplasm-Localized PP2A B' Subunits.Mol Plant. 2016 Jan 4;9(1):148-157. doi: 10.1016/j.molp.2015.10.007. Epub 2015 Oct 27. Mol Plant. 2016. PMID: 26517938
-
Emerging Roles of B56 Phosphorylation and Binding Motif in PP2A-B56 Holoenzyme Biological Function.Int J Mol Sci. 2024 Mar 10;25(6):3185. doi: 10.3390/ijms25063185. Int J Mol Sci. 2024. PMID: 38542160 Free PMC article. Review.
-
Distinct Clades of Protein Phosphatase 2A Regulatory B'/B56 Subunits Engage in Different Physiological Processes.Int J Mol Sci. 2023 Jul 31;24(15):12255. doi: 10.3390/ijms241512255. Int J Mol Sci. 2023. PMID: 37569631 Free PMC article. Review.
Cited by
-
A complex of BRCA2 and PP2A-B56 is required for DNA repair by homologous recombination.Nat Commun. 2021 Sep 30;12(1):5748. doi: 10.1038/s41467-021-26079-0. Nat Commun. 2021. PMID: 34593815 Free PMC article.
-
Micropeptide CIP2A-BP encoded by LINC00665 inhibits triple-negative breast cancer progression.EMBO J. 2020 Jan 2;39(1):e102190. doi: 10.15252/embj.2019102190. Epub 2019 Nov 22. EMBO J. 2020. PMID: 31755573 Free PMC article.
-
Reduction of protein phosphatase 2A (PP2A) complexity reveals cellular functions and dephosphorylation motifs of the PP2A/B'δ holoenzyme.J Biol Chem. 2020 Apr 24;295(17):5654-5668. doi: 10.1074/jbc.RA119.011270. Epub 2020 Mar 10. J Biol Chem. 2020. PMID: 32156701 Free PMC article.
-
The Expression Regulation and Cancer-Promoting Roles of RACGAP1.Biomolecules. 2024 Dec 24;15(1):3. doi: 10.3390/biom15010003. Biomolecules. 2024. PMID: 39858398 Free PMC article. Review.
-
Structural insights into PPP2R5A degradation by HIV-1 Vif.Nat Struct Mol Biol. 2024 Oct;31(10):1492-1501. doi: 10.1038/s41594-024-01314-6. Epub 2024 May 24. Nat Struct Mol Biol. 2024. PMID: 38789685 Free PMC article.
References
-
- Ubersax JA, Ferrell JE Jr. Mechanisms of specificity in protein phosphorylation. Nat Rev Mol Cell Biol 2007; 8: 530–541. - PubMed
-
- Daub H, Olsen JV, Bairlein M et al. Kinase-selective enrichment enables quantitative phosphoproteomics of the kinome across the cell cycle. Mol Cell 2008; 31: 438–448. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
