

CDK10 Mutations in Humans and Mice Cause Severe Growth Retardation, Spine Malformations, and Developmental Delays
- PMID: 28886341
- PMCID: PMC5591019
- DOI: 10.1016/j.ajhg.2017.08.003
CDK10 Mutations in Humans and Mice Cause Severe Growth Retardation, Spine Malformations, and Developmental Delays
Abstract
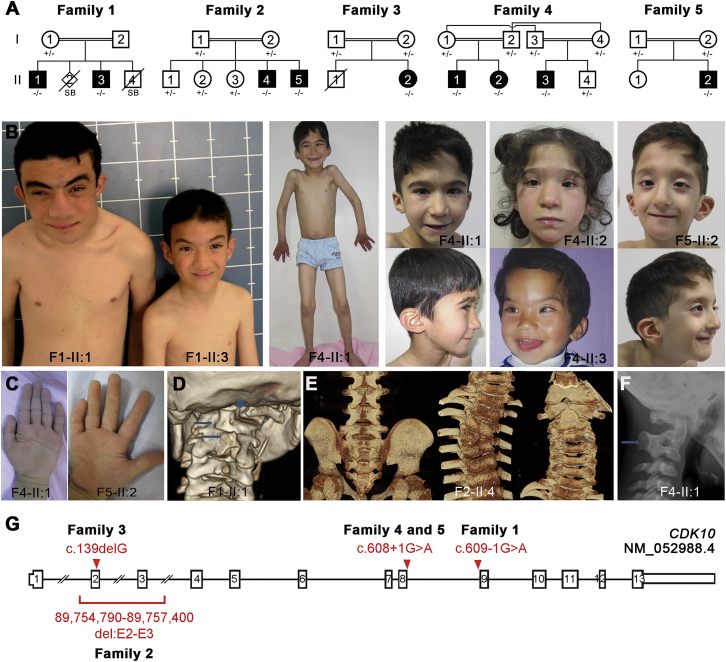
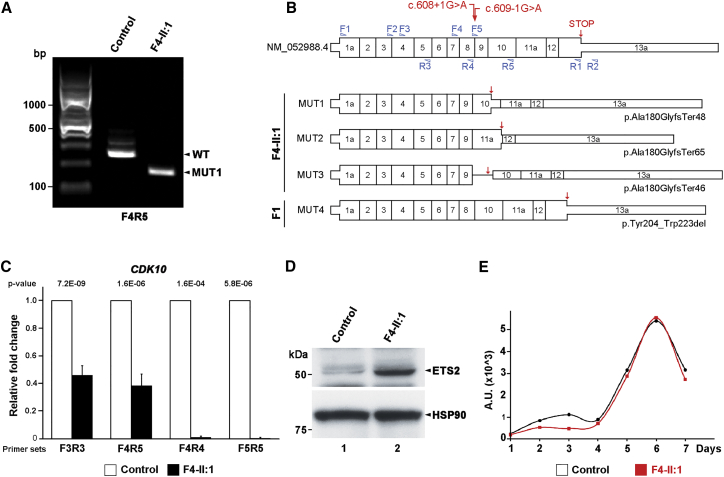
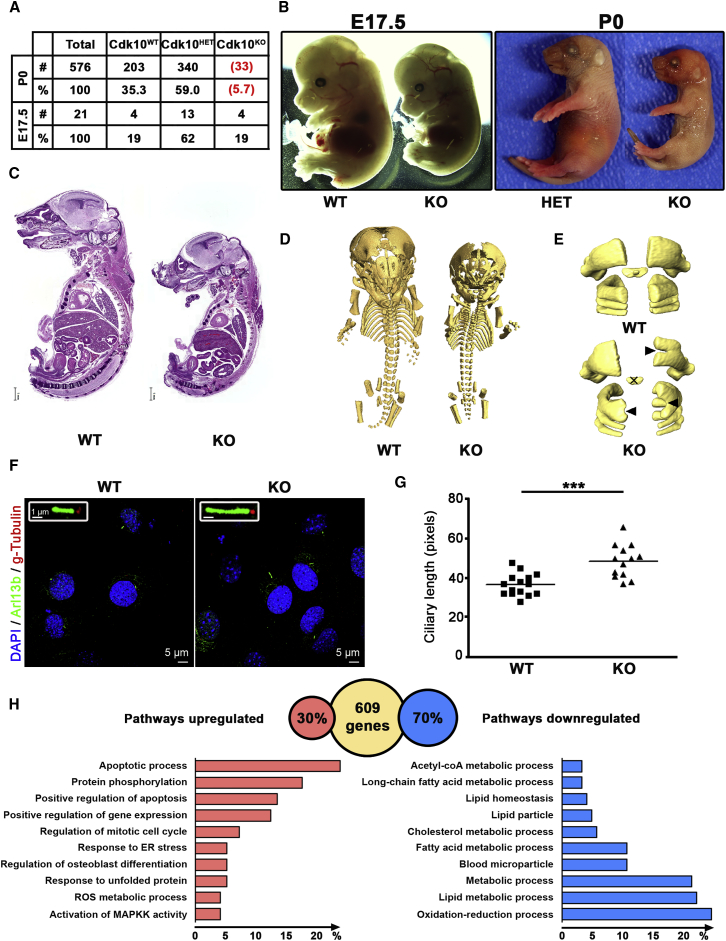
In five separate families, we identified nine individuals affected by a previously unidentified syndrome characterized by growth retardation, spine malformation, facial dysmorphisms, and developmental delays. Using homozygosity mapping, array CGH, and exome sequencing, we uncovered bi-allelic loss-of-function CDK10 mutations segregating with this disease. CDK10 is a protein kinase that partners with cyclin M to phosphorylate substrates such as ETS2 and PKN2 in order to modulate cellular growth. To validate and model the pathogenicity of these CDK10 germline mutations, we generated conditional-knockout mice. Homozygous Cdk10-knockout mice died postnatally with severe growth retardation, skeletal defects, and kidney and lung abnormalities, symptoms that partly resemble the disease's effect in humans. Fibroblasts derived from affected individuals and Cdk10-knockout mouse embryonic fibroblasts (MEFs) proliferated normally; however, Cdk10-knockout MEFs developed longer cilia. Comparative transcriptomic analysis of mutant and wild-type mouse organs revealed lipid metabolic changes consistent with growth impairment and altered ciliogenesis in the absence of CDK10. Our results document the CDK10 loss-of-function phenotype and point to a function for CDK10 in transducing signals received at the primary cilia to sustain embryonic and postnatal development.
Keywords: Al Kaissi syndrome knockout mice; CDK10; ETS2; cilia; congenital disorder; growth retardation; metabolism; spine malformation.
Copyright © 2017 American Society of Human Genetics. All rights reserved.
Figures
References
-
- Lim S., Kaldis P. Cdks, cyclins and CKIs: roles beyond cell cycle regulation. Development. 2013;140:3079–3093. - PubMed
-
- Morgan D.O. New Science Press Ltd; 2007. The cell cycle: principles of control.
-
- Brambilla R., Draetta G. Molecular cloning of PISSLRE, a novel putative member of the cdk family of protein serine/threonine kinases. Oncogene. 1994;9:3037–3041. - PubMed
-
- Graña X., Claudio P.P., De Luca A., Sang N., Giordano A. PISSLRE, a human novel CDC2-related protein kinase. Oncogene. 1994;9:2097–2103. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
